De geslachtsbepaling het wordt gecontroleerd door een reeks zeer uiteenlopende mechanismen onder taxa, die de geslachtskenmerken van het individu vaststellen. Deze systemen kunnen intrinsiek zijn voor het individu - dat wil zeggen genetisch - of worden beheerst door de omgevingsfactoren die het individu omringen tijdens de vroege stadia van zijn leven..
Bij intrinsieke bepaling hebben biologen deze systemen in drie hoofdcategorieën ingedeeld: individuele genen, haplodiploïde systeem of speciale of geslachtschromosomen. Het laatste geval is dat van ons, zoogdieren, vogels en sommige insecten.

Op dezelfde manier beïnvloeden omgevingsfactoren ook de geslachtsbepaling. Dit fenomeen is bestudeerd bij sommige reptielen en amfibieën, die vooral door temperatuur worden beïnvloed. Dit vaststellingssysteem staat bekend als cryptisch.
Artikel index
Seks, begrepen als het mengen van genomen via meiose en gametenfusie, is een vrijwel universele gebeurtenis in het leven van eukaryoten..
Een van de belangrijkste gevolgen van seksuele voortplanting is de koppeling van verschillende allelen, gedragen door verschillende individuen, in een gunstige genetische variatie..
Bij de meeste eukaryote organismen is geslachtsbepaling een gebeurtenis die plaatsvindt op het moment van bevruchting. Dit fenomeen kan optreden door drie verschillende systemen: individuele genen, haplodiploïde systeem of speciale chromosomen..
Evenzo hebben we de bepaling van geslachtskenmerken gemedieerd door omgevingsfactoren, zoals temperatuur. Dit komt voor bij kikkers, schildpadden en alligators, waar de incubatietemperaturen het geslacht lijken te bepalen..
We zullen elk systeem hieronder beschrijven aan de hand van voorbeelden uit de dieren- en plantenrijken:

In organismen waar het geslacht wordt bepaald door individuele genen, zijn er geen geslachtschromosomen. In deze gevallen hangt het geslacht af van een reeks allelen die zich op specifieke chromosomen bevinden..
Met andere woorden, geslacht wordt bepaald door een gen (of door meerdere hiervan) en niet door de aanwezigheid van een volledig chromosoom.
Verschillende gewervelde dieren, zoals vissen, amfibieën en sommige reptielen, hebben dit systeem. Het is ook gemeld bij planten.
De allelen die deelnemen aan dit fenomeen hebben het algemeen bekende dominantiesysteem dat bestaat voor autosomale karakters. In planten zijn de allelen gespecificeerd die mannelijkheid, hermafroditisme en het vrouwelijke karakter van het individu bepalen..

Haplodiploïde systemen bepalen het geslacht afhankelijk van de haploïde of diploïde toestand van het individu. Wij mensen zijn diploïde - zowel mannen als vrouwen. Deze voorwaarde kan echter niet naar alle diergroepen worden geëxtrapoleerd..
Het haplodiploïde systeem komt vrij veel voor bij Hymenoptera (bijen, mieren en dergelijke), Homoptera (wolluizen en kuikens) en Coleoptera (kevers).
Het klassieke voorbeeld is dat van bijen en de geslachtsbepaling in kolonies. De sociale structuur van bijen is buitengewoon complex, net als hun eusociaal gedrag, met hun basis in het genetische systeem dat hun geslacht bepaalt.
Bijen hebben geen geslachtschromosomen. Vrouwtjes zijn diploïde (2n) en mannetjes zijn haploïde (n), de zogenaamde drones. Daarom komt de ontwikkeling van vrouwtjes voort uit de bevruchting van de eieren, terwijl de onbevruchte eitjes zich ontwikkelen tot mannetjes. Dat wil zeggen, de laatsten hebben geen vader.
Bij vrouwen is de scheiding tussen de werksters en de koningin niet genetisch bepaald. Deze hiërarchie wordt bepaald door het dieet van het individu in de vroege stadia van zijn leven..
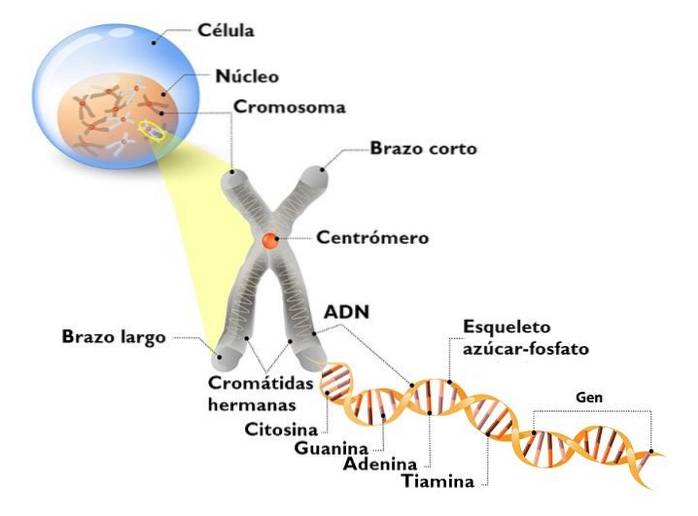
Het geval van speciale chromosomen of geslachtschromosomen is het geval waarmee we het meest verwant zijn. Het is aanwezig in alle zoogdieren, alle vogels en veel insecten, en is een veel voorkomende vorm in organismen met verschillende seksuele fenotypes.
In planten, hoewel het zeer zeldzaam is, zijn sommige dioccale soorten gespecificeerd die geslachtschromosomen hebben..
Dit systeem kent verschillende varianten. Onder de meest voorkomende en eenvoudigste vinden we de systemen: XX-X0 en XX-XY, waar het heterogametische geslacht het mannetje is, en de ZZ-ZW, waar het heterogametische geslacht het vrouwtje is..
Het eerste systeem, XX en X0, komt veel voor bij insecten van de orde Orthoptera en Hemiptera. In deze gevallen heeft het mannetje maar één geslachtschromosoom.
Het XX- en XY-systeem is aanwezig bij zoogdieren, bij veel insecten van de Diptera-orde en in een zeer beperkt aantal planten, zoals Cannabis sativa. In dit systeem wordt geslacht bepaald door de mannelijke gameet. Als de laatste het X-chromosoom heeft, komt het nageslacht overeen met een vrouw, terwijl de Y-gameet aanleiding geeft tot een mannetje.
Het laatste systeem, ZZ en ZW, is aanwezig bij alle vogels en bij sommige insecten van de orde Lepidoptera

In bepaalde taxa spelen de verschillende omgevingsstimuli, in de eerste levensfasen van de individuen, een cruciale rol bij de bepaling van het geslacht. In deze gevallen is de bepaling vanuit genetisch oogpunt niet volledig opgehelderd en lijkt het geslacht volledig afhankelijk te zijn van de omgeving..
Bij zeeschildpadden bijvoorbeeld, verandert een variatie van 1 ° C extra een hele populatie mannetjes in een populatie die uitsluitend uit vrouwtjes bestaat..
Bij alligators is gevonden dat een incubatie onder 32 ° C een populatie vrouwtjes oplevert en temperaturen boven 34 ° C vertaalt zich in een populatie mannetjes. In het bereik van 32 tot 34 zijn de verhoudingen tussen de geslachten variabel.
Naast temperatuur is de invloed van andere omgevingsvariabelen aangetoond. In een soort ringworm, Bonellia viridis, geslacht wordt bepaald in het larvale stadium. De larven die vrij in het water zwemmen, ontwikkelen zich als mannetjes.
Daarentegen worden de larven die zich ontwikkelen tot bijna volwassen vrouwtjes, omgezet in mannetjes, door bepaalde hormonen die deze afscheiden.
Ten slotte bespreken we het speciale geval van hoe de aanwezigheid van een bacterie het geslacht van een populatie kan bepalen. Dit is het geval bij de beroemde bacteriën die tot het geslacht behoren Wolbachia.
Wolbachia het is een intracellulaire symbiont, die in staat is om een breed scala aan geleedpotige soorten en ook enkele nematoden te infecteren. Deze bacterie wordt verticaal overgedragen, van vrouwtjes op hun toekomstige nakomelingen, door eieren - hoewel ook horizontale overdracht is gedocumenteerd..
Met betrekking tot de bepaling van het geslacht in de organismen die er wonen, Wolbachia heeft zeer relevante effecten.
Het is in staat mannetjes in de populatie te doden, waarbij geïnfecteerde mannetjes sterven tijdens de vroege stadia van hun leven; feminiseert de bevolking, waar zich ontwikkelende mannetjes in vrouwtjes veranderen; en tenslotte is het in staat parthenogenetische populaties te produceren.
Al deze genoemde fenotypen, die de vervorming van de geslachtsverhouding met zich meebrengen met een duidelijke voorkeur voor vrouwen, treden op om de overdracht van de bacteriën naar de volgende generatie te bevorderen..
Dankzij het brede scala aan hosts, Wolbachia heeft een cruciale rol gespeeld in de evolutie van de systemen voor geslachtsbepaling en reproductiestrategieën van geleedpotigen.

Een fundamentele eigenschap van systemen voor geslachtsbepaling is het begrijpen van de verhouding tussen de seksen of geslachtsverhouding. Er zijn verschillende theorieën en hypothesen voorgesteld:
Ronald Fisher, een veelgeprezen Britse statisticus en bioloog, stelde in 1930 een theorie voor om uit te leggen waarom populaties een verhouding van 50:50 tussen mannen en vrouwen handhaven. Redelijkerwijs verklaarde het ook waarom de mechanismen die deze gelijke verhouding scheeftrekken, worden geselecteerd tegen.
Achtereenvolgens was het mogelijk om aan te tonen dat een eerlijke of evenwichtige geslachtsverhouding een stabiele strategie vormt, vanuit evolutionair oogpunt.
Het is waar dat de resultaten van Fisher in bepaalde omstandigheden niet van toepassing zijn, maar zijn hypothese lijkt algemeen genoeg te zijn dat de mechanismen voor het bepalen van het geslacht volgens zijn principes moeten worden gekozen..
Later, in 1973, merkten deze auteurs op dat de geslachtsverhouding afhankelijk was van vele andere factoren - voornamelijk de fysiologische toestand van de vrouw - waarmee in de verklaring van Fisher geen rekening werd gehouden..
Het argument was gebaseerd op de volgende premissen: wanneer een vrouwtje fysiologisch "gezond" is, zou ze mannetjes moeten produceren, omdat deze kleintjes een grotere kans hebben om te overleven en zich voort te planten..
Op dezelfde manier, wanneer het vrouwtje niet in optimale fysiologische omstandigheden verkeert, is de beste strategie de productie van andere vrouwtjes..
In de natuur planten zwakke vrouwtjes zich vaak voort, ondanks hun fysiologische "minderwaardigheids" -status. In tegenstelling tot een zwakke man, waar de kans op voortplanting uitzonderlijk lager is.
Dit voorstel is getest in verschillende biologische systemen, zoals ratten, herten, zeehonden en zelfs in menselijke populaties..
In het licht van de evolutie roept de diversiteit van de mechanismen die het geslacht bepalen bepaalde vragen op, waaronder: waarom zien we deze variatie, hoe ontstaat deze variatie en ten slotte, waarom treden deze veranderingen op??
Bovendien komt het ook voort uit de vraag of bepaalde mechanismen het individu een bepaald voordeel geven ten opzichte van andere. Dat wil zeggen als een bepaald mechanisme selectief de voorkeur heeft gekregen.



Niemand heeft nog op dit artikel gereageerd.