De erytropoëtine, hemopoëtine of EPO is een glycoproteïne met hormonale functies (cytokine) dat verantwoordelijk is voor de controle van de proliferatie, differentiatie en overleving van de voorlopercellen van erytrocyten of rode bloedcellen in het beenmerg, dat wil zeggen van erytropoëse.
Dit eiwit is een van de verschillende groeifactoren die de hematopoëtische processen regelen waardoor uit een kleine groep pluripotente stamcellen de in het bloed gevonden cellen worden gevormd: zowel erytrocyten als witte bloedcellen en lymfocyten. Dat wil zeggen, cellen van de myeloïde en lymfoïde lijnen.
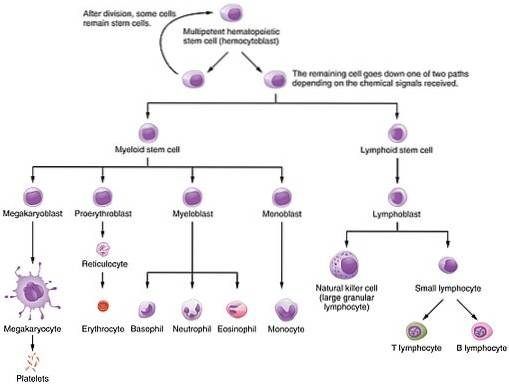
Het belang ervan ligt in het functionele belang van cellen die helpen zich te vermenigvuldigen, differentiëren en rijpen, aangezien erytrocyten verantwoordelijk zijn voor het transport van zuurstof van de longen naar de verschillende weefsels van het lichaam..
Erytropoëtine was de eerste groeifactor die werd gekloond (in 1985), en de toediening ervan voor de succesvolle behandeling van bloedarmoede veroorzaakt door nierfalen is momenteel goedgekeurd door de Amerikaanse Food and Drug Administration (FDA)..
Het idee dat erytropoëse wordt beheerst door een humorale factor (oplosbare factor aanwezig in de bloedsomloop) werd meer dan 100 jaar geleden voorgesteld door Carnot en Deflandre bij het bestuderen van de positieve effecten op de toename van het percentage rode bloedcellen bij konijnen die met serum werden behandeld. dieren.
Pas in 1948 introduceerden Bonsdorff en Jalavisto de term "erytropoëtine" om de humorale factor te beschrijven met een specifieke implicatie voor de productie van erytrocyten..
Artikel index
Erytropoëtine is een eiwit uit de glycoproteïnefamilie. Het is stabiel bij zure pH's en heeft een molecuulgewicht van ongeveer 34 kDa..
Het heeft ongeveer 193 aminozuren, waaronder een hydrofoob N-terminaal gebied met 27 resten, dat wordt verwijderd door co-translationele verwerking; en een arginineresidu op positie 166 dat ook verloren gaat, dus het circulerende eiwit heeft 165 aminozuren.
In zijn structuur is de vorming van twee disulfidebruggen tussen de cysteïneresiduen aanwezig op de posities 7-161 en 29-33 te zien, die verband houden met de werking ervan. Het bestaat voor meer of minder dan 50% uit alfa-helices, die blijkbaar deelnemen aan de vorming van een bolvormig gebied of deel.
Het heeft 40% koolhydraten, vertegenwoordigd door drie oligosaccharideketens N-gebonden aan verschillende asparaginezuurresiduen (Asp) en een O-keten gekoppeld aan een serineresidu (Ser). Deze oligosacchariden zijn voornamelijk samengesteld uit fucose, mannose, N-acetylglucosamine, galactose en N-acetylneuraminezuur..
De koolhydraatregio van EPO vervult verschillende rollen:
- Het is essentieel voor zijn biologische activiteit.
- Beschermt het tegen afbraak of schade veroorzaakt door vrije zuurstofradicalen.
- De oligosaccharideketens zijn vereist voor de uitscheiding van het rijpe eiwit.
Bij mensen bevindt het gen dat codeert voor dit eiwit zich in het midden van de lange arm van chromosoom 7, in het gebied q11-q22; het wordt gevonden in een enkele kopie in een gebied van 5,4 kb en heeft vijf exons en vier introns. Homologiestudies geven aan dat de sequentie ervan 92% gelijkenis vertoont met die van andere primaten en 80% met die van sommige knaagdieren..
Tijdens de ontwikkeling van de foetus wordt erytropoëtine voornamelijk in de lever geproduceerd, maar er is vastgesteld dat in dezelfde fase het gen dat codeert voor dit hormoon ook overvloedig tot expressie komt in het middengebied van de niernefronen..
Na de geboorte, in wat als alle postnatale stadia kan worden beschouwd, wordt het hormoon voornamelijk in de nieren geproduceerd. Specifiek door de cellen van de cortex en het oppervlak van de nierlichaampjes.
De lever neemt ook deel aan de productie van erytropoëtine in de postnatale stadia, waaruit ongeveer 20% van het totale circulerende EPO-gehalte wordt uitgescheiden..
Andere "extra-renale" organen waar de productie van erytropoëtine is gedetecteerd, zijn onder meer perifere endotheelcellen, vasculaire gladde spiercellen en insulineproducerende cellen..
Van sommige EPO-secretiecentra is ook bekend dat ze voorkomen in het centrale zenuwstelsel, inclusief de hippocampus, cortex, hersenendotheelcellen en astrocyten..
De productie van erytropoëtine wordt niet rechtstreeks gestuurd door het aantal rode bloedcellen in het bloed, maar door de toevoer van zuurstof in de weefsels. Een tekort aan zuurstof in de weefsels stimuleert de productie van EPO en zijn receptoren in de lever en de nieren.
Deze door hypoxie gemedieerde activering van genexpressie is het product van de activering van de route van een familie van transcriptiefactoren die bekend staat als hypoxie-induceerbare factor 1 (HIF-1). hypoxie-induceerbare factor 1.
Hypoxie induceert dan de vorming van veel eiwitcomplexen die verschillende functies vervullen bij de activering van erytropoëtine-expressie, en die direct of indirect binden aan factoren die het activeringssignaal vertalen naar de promotor van het EPO-gen, waardoor de transcriptie ervan wordt gestimuleerd..
Andere stressfactoren zoals hypoglykemie (lage bloedsuikerspiegel), verhogingen van intracellulair calcium of de aanwezigheid van reactieve zuurstofsoorten, triggeren ook de HIF-1-route..
Het werkingsmechanisme van erytropoëtine is vrij complex en hangt voornamelijk af van het vermogen om verschillende signaalcascades te stimuleren die betrokken zijn bij celproliferatie, die op hun beurt verband houden met de activering van andere factoren en hormonen..
In het menselijk lichaam van een gezonde volwassene is er een evenwicht tussen de productie en vernietiging van rode bloedcellen of erytrocyten, en EPO draagt bij aan het handhaven van dit evenwicht door de verdwijnende erytrocyten te vervangen..
Wanneer de hoeveelheid zuurstof die beschikbaar is in de weefsels erg laag is, neemt de expressie van het gen dat codeert voor erytropoëtine toe in de nieren en de lever. De prikkel kan ook worden gegeven door grote hoogten, hemolyse, aandoeningen van ernstige bloedarmoede, bloeding of langdurige blootstelling aan koolmonoxide.
Deze omstandigheden veroorzaken een toestand van hypoxie, waardoor de afscheiding van EPO toeneemt, een groter aantal rode bloedcellen wordt geproduceerd en ook de fractie reticulocyten in circulatie, die een van de voorlopercellen van erytrocyten zijn, toeneemt..
Bij erytropoëse is EPO voornamelijk betrokken bij de proliferatie en differentiatie van voorlopercellen die betrokken zijn bij de afstamming van rode bloedcellen (erythrocytische voorlopercellen), maar het activeert ook mitose bij pro-erythroblasten en basofiele erytroblasten, en versnelt ook de afgifte van reticulocyten uit het beenmerg..
Het eerste niveau waarop het eiwit werkt, is het voorkomen van geprogrammeerde celdood (apoptose) van de voorlopercellen die in het beenmerg worden gevormd, wat het bereikt door remmende interactie met de factoren die bij dit proces betrokken zijn..
Cellen die reageren op erytropoëtine bezitten een specifieke receptor voor erytropoëtine, bekend als de erytropoëtinereceptor of EpoR. Zodra het eiwit een complex vormt met zijn receptor, wordt het signaal overgedragen in de cel: naar de celkern.
De eerste stap voor signaaloverdracht is een conformatieverandering die optreedt nadat het eiwit bindt aan zijn receptor, die tegelijkertijd is gebonden aan andere receptormoleculen die worden geactiveerd. Onder hen is Janus-tyrosine kinase 2 (Jack-2).
Onder enkele van de routes die stroomafwaarts worden geactiveerd, nadat Jack-2 de fosforylering van tyrosineresiduen van de EpoR-receptor bemiddelt, is de MAP-kinase- en proteïnekinase C-route, die transcriptiefactoren activeren die de expressie van specifieke genen verhogen..
Zoals veel hormonale factoren in organismen, is erytropoëtine niet beperkt tot een enkele functie. Dit is door middel van talrijke onderzoeken opgehelderd.
Naast het fungeren als een proliferatie- en differentiatiefactor voor erytrocyten, die essentieel zijn voor het transport van gassen door de bloedbaan, lijkt erytropoëtine enkele aanvullende functies te vervullen die niet noodzakelijk verband houden met de activering van celproliferatie en differentiatie..
Studies hebben gesuggereerd dat EPO celbeschadiging voorkomt en, hoewel de werkingsmechanismen ervan niet precies bekend zijn, wordt aangenomen dat het apoptotische processen kan voorkomen die worden geproduceerd door verminderde of afwezige zuurstofspanning, exciteertoxiciteit en blootstelling aan vrije radicalen..
Zijn deelname aan de preventie van apoptose is onderzocht door interactie met bepalende factoren in de signaalcascades: Janus-tyrosinekinase 2 (Jak2), caspase 9, caspase 1 en caspase 3, glycogeensynthase-kinase-3β, activeringsfactor van apoptotische proteasen 1 (Apaf-1) en anderen.
Neemt deel aan de remming van cellulaire ontsteking door enkele pro-inflammatoire cytokines te remmen, zoals interleukine 6 (IL-6), tumornecrosefactor alfa (TNF-α) en monocyt chemo-lokstofproteïne 1.
In het vasculaire systeem is aangetoond dat het samenwerkt bij het handhaven van de integriteit en bij de vorming van nieuwe capillairen uit bestaande vaten in gebieden zonder vasculatuur (angiogenese). Bovendien voorkomt het de doorlaatbaarheid van de bloed-hersenbarrière tijdens blessures.
Aangenomen wordt dat het postnatale neovascularisatie stimuleert door de mobilisatie van voorlopercellen van het beenmerg naar de rest van het lichaam te vergroten..
Het speelt een belangrijke rol bij de ontwikkeling van neurale voorlopercellen door de activering van de nucleaire factor KB, die de productie van zenuwstamcellen bevordert..
EPO werkt samen met andere cytokinen en heeft een "modulerende" functie bij de controle van proliferatie- en differentiatieroutes van megakaryocyten en granulocyt-monocyten..



Niemand heeft nog op dit artikel gereageerd.