De DNA-verpakking is een term die de gecontroleerde verdichting van DNA in de cel definieert. In geen enkele cel (en zelfs niet in virussen) is DNA vrij, los en in echte oplossing.
DNA is een extreem lang molecuul dat ook altijd in wisselwerking staat met een grote verscheidenheid aan verschillende eiwitten. Voor de verwerking, overerving en controle van de expressie van de genen die het draagt, neemt DNA een bepaalde ruimtelijke organisatie aan. Dit wordt bereikt door de cel door elke stap van het verpakken van DNA strikt te controleren op verschillende verdichtingsniveaus..
Virussen hebben verschillende verpakkingsstrategieën voor hun nucleïnezuren. Een van de favorieten is die van de vorming van compacte spiralen. Je zou kunnen zeggen dat virussen nucleïnezuren zijn die verpakt zijn in hun eigen eiwitten die ze bedekken, beschermen en mobiliseren.
In prokaryoten wordt DNA geassocieerd met eiwitten die de vorming van complexe lussen in een structuur genaamd nucleoïde bepalen. Het maximale niveau van DNA-verdichting in een eukaryote cel is daarentegen het mitotische of meiotische chromosoom..
Het enige geval waarin een B-DNA niet is verpakt, is een onderzoekslaboratorium dat dit doel nastreeft..
Artikel index
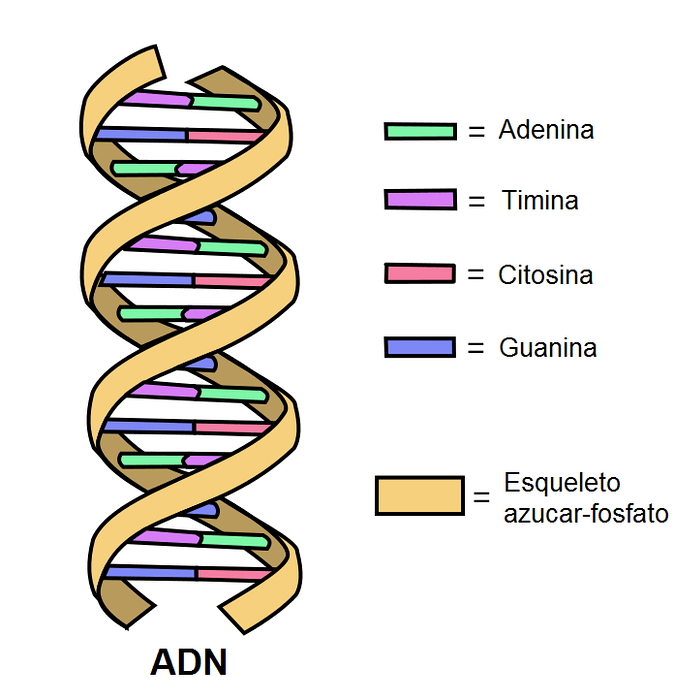
DNA bestaat uit twee antiparallelle banden die een dubbele helix vormen. Elk van hen heeft een fosfodiësterbindingsskelet waarop suikers die zijn gekoppeld aan stikstofbasen, zijn bevestigd..
Binnen het molecuul vormen de stikstofbasen van één band waterstofbruggen (twee of drie) met de complementaire band.
In een molecuul als dit vertonen de meeste belangrijke bindingshoeken vrije rotatie. Stikstof-basissuiker-, suiker-fosfaatgroep- en fosfodiësterbindingen zijn flexibel.
Hierdoor kan DNA, gezien als een flexibele staaf, enig vermogen vertonen om te buigen en te draaien. Deze flexibiliteit stelt DNA in staat om complexe lokale structuren aan te nemen en interactielussen te vormen op korte, middellange en lange afstanden..
Deze flexibiliteit legt ook uit hoe 2 meter DNA kan worden vastgehouden in elke diploïde cel van een mens. In een gameet (haploïde cel) zou het een meter DNA zijn.
Hoewel het geen onbreekbare regel is, bestaat het bacteriële chromosoom als een enkelvoudig supergewonden dubbelbandig DNA-molecuul..
De dubbele helix draait meer op zichzelf (meer dan 10 bp per omwenteling) en produceert dus enige verdichting. Lokale knopen worden ook gegenereerd dankzij manipulaties die enzymatisch worden gecontroleerd.
Bovendien zijn er sequenties in DNA die het mogelijk maken dat domeinen zich in grote lussen vormen. We noemen de structuur die het resultaat is van de supercoil en geordende lussen een nucleoïde..
Deze ondergaan dynamische veranderingen dankzij enkele eiwitten die enige structurele stabiliteit bieden aan het verdichte chromosoom. De mate van verdichting bij bacteriën en archaea is zo efficiënt dat er meer dan één chromosoom per nucleoïde kan zijn.
De nucleoïde comprimeert het prokaryotische DNA minstens 1000 keer. De zeer topologische structuur van de nucleoïde is een fundamenteel onderdeel van de regulatie van de genen die door het chromosoom worden gedragen. Dat wil zeggen, structuur en functie vormen dezelfde eenheid..
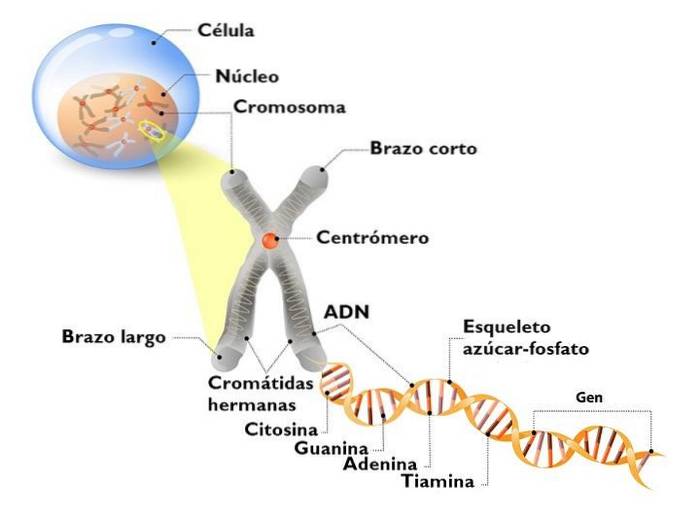
Het DNA in de eukaryote kern is niet naakt. Het interageert met veel eiwitten, waarvan de belangrijkste histonen zijn. Histonen zijn kleine, positief geladen eiwitten die niet-specifiek aan DNA binden.
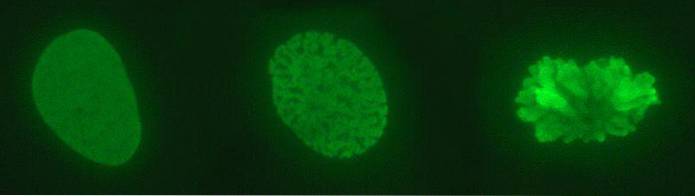
In de kern zien we een complex DNA: histonen, die we chromatine noemen. Het sterk gecondenseerde chromatine, dat over het algemeen niet tot expressie wordt gebracht, is heterochromatine. Daarentegen is de minst verdichte (lossere) of euchromatine chromatine met genen die tot expressie worden gebracht.
Chromatine heeft verschillende verdichtingsniveaus. Het meest elementaire is dat van het nucleosoom; Het wordt gevolgd door de solenoïde vezel en interfase chromatinelussen. Alleen wanneer een chromosoom zich deelt, worden maximale verdichtingsniveaus weergegeven..
Het nucleosoom is de basiseenheid van de organisatie van chromatine. Elk nucleosoom bestaat uit een octameer van histonen die een soort trommel vormen..
De octameer bestaat uit twee kopieën van elk van de histonen H2A, H2B, H3 en H4. Om hen heen gaat het DNA ongeveer 1,7 keer. Het wordt gevolgd door een fractie van vrij DNA, een 20 bp-linker genaamd, geassocieerd met histon H1, en vervolgens een ander nucleosoom. De hoeveelheid DNA in een nucleosoom en dat wat het aan een ander bindt, is ongeveer 166 basenparen.
Deze stap van het inpakken van DNA comprimeert het molecuul ongeveer 7 keer. Dat wil zeggen, we gaan van een meter naar iets meer dan 14 cm DNA.
Deze pakking is mogelijk omdat de positieve histonen de negatieve lading van het DNA en de daaruit voortvloeiende elektrostatische zelfafstoting opheffen. De andere reden is dat DNA zo kan worden gevouwen dat het de octameer van histonen kan omdraaien..
De vezel van kralen in een halsketting gevormd door vele opeenvolgende nucleosomen wordt verder gewikkeld tot een meer verdichte structuur.
Hoewel we niet duidelijk zijn welke structuur het daadwerkelijk aanneemt, weten we wel dat het een dikte bereikt van ongeveer 30 nm. Dit is de zogenaamde 30 nm-vezel; Histone H1 is essentieel voor zijn vorming en stabiliteit.
De 30 nm-vezel is de structurele basiseenheid van heterochromatine. Dat van losse nucleosomen, dat van euchromatine.
De 30 nm-vezel is echter niet volledig lineair. Integendeel, het vormt lussen van ongeveer 300 nm lang, op een kronkelige manier, op een weinig bekende eiwitmatrix..
Deze lussen op een eiwitmatrix vormen een compactere chromatinevezel met een diameter van 250 nm. Ten slotte worden ze uitgelijnd als een enkele helix met een dikte van 700 nm, wat aanleiding geeft tot een van de zusterchromatiden van een mitotisch chromosoom..
Uiteindelijk comprimeert het DNA in nucleair chromatine ongeveer 10.000 keer op het chromosoom van de delende cel. In de interfase-kern is de verdichting ook hoog, aangezien het ongeveer 1000 keer is vergeleken met het "lineaire" DNA..

In de wereld van ontwikkelingsbiologie zou gametogenese het epigenoom resetten. Dat wil zeggen, het wist de DNA-markeringen die het leven van de persoon die aanleiding gaf tot de gameet, produceerde of ervoer.
Deze tags omvatten DNA-methylatie en covalente modificaties van histonen (code voor histonen). Maar niet het hele epigenoom wordt gereset. Wat achterblijft met merktekens is verantwoordelijk voor de vaderlijke of moederlijke genetische afdruk.
De impliciete reset naar gametogenese is gemakkelijker te zien in sperma. In sperma zit DNA niet vol met histonen. Daarom wordt de informatie die verband houdt met de wijzigingen ervan in het producerende organisme in het algemeen niet geërfd..
In sperma wordt DNA verpakt door interactie met niet-specifieke DNA-bindende eiwitten die protamines worden genoemd. Deze eiwitten vormen disulfidebindingen met elkaar, waardoor overlappende DNA-lagen worden gevormd die niet elektrostatisch afstoten..



Niemand heeft nog op dit artikel gereageerd.