De homologe recombinatie het is een proces waarbij DNA-moleculen worden uitgewisseld tussen vergelijkbare of identieke delen van het genoom. Cellen gebruiken homologe recombinatie voornamelijk om breuken in genetisch materiaal te herstellen, waardoor genetische variatie in populaties ontstaat.
In het algemeen omvat homologe recombinatie de fysieke koppeling tussen homologe gebieden van het genetisch materiaal, gevolgd door het verbreken van de ketens die de uitwisseling zullen ondergaan, en tenslotte de vereniging van de nieuwe gecombineerde DNA-moleculen..
Breuken in het DNA moeten zo snel en efficiënt mogelijk worden hersteld. Als de schade niet wordt hersteld, kunnen de gevolgen ernstig en zelfs dodelijk zijn. Bij bacteriën is de belangrijkste functie van homologe recombinatie het herstellen van deze breuken in het genetisch materiaal.
Homologe recombinatie wordt beschouwd als een van de belangrijkste mechanismen die de stabiliteit van het genoom mogelijk maken. Het is aanwezig in alle domeinen van het leven en zelfs in virussen, dus het is vermoedelijk een vitaal mechanisme dat al heel vroeg in de evolutie van het leven verscheen..
Artikel index
Een van de meest relevante principes die door Gregor Mendel wordt voorgesteld, is onafhankelijkheid in de scheiding van karakters. Volgens deze wet worden de verschillende genen onafhankelijk van ouder op kind overgedragen.
In 1900 waren er echter duidelijke uitzonderingen op dit principe. De Engelse genetici Bateson en Punnett toonden aan dat bepaalde karakters vaak samen worden overgeërfd, en voor deze eigenschappen heeft het principe dat door Mendel wordt verkondigd geen geldigheid..
Later onderzoek slaagde erin het bestaan van het recombinatieproces op te helderen, waarbij cellen in staat waren genetisch materiaal uit te wisselen. In gevallen waarin de genen samen erfden, werd het DNA niet uitgewisseld vanwege de fysieke nabijheid tussen de genen..
Homologe recombinatie is een cellulair fenomeen waarbij de fysieke uitwisseling van DNA-sequenties tussen twee chromosomen betrokken is. Bij recombinatie is een reeks genen betrokken die bekend staan als genen rec. Deze coderen voor verschillende enzymen die aan het proces deelnemen.
DNA-moleculen worden als "homoloog" beschouwd wanneer ze vergelijkbare of identieke sequenties van meer dan 100 basenparen delen. DNA heeft kleine regio's die van elkaar kunnen verschillen, en deze varianten staan bekend als allelen.
In levende wezens wordt al het DNA als recombinant DNA beschouwd. De uitwisseling van genetisch materiaal tussen chromosomen vindt continu plaats, waarbij de genen op de chromosomen worden gemengd en herschikt.
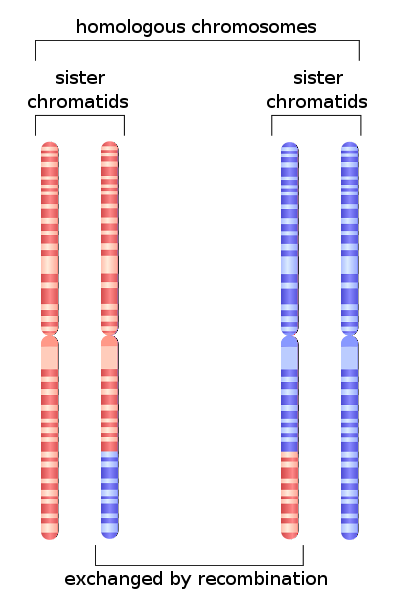
Dit proces vindt duidelijk plaats in meiose. Specifiek in de fase waarin chromosomen paren in de eerste celdeling. In deze fase vindt de uitwisseling van genetisch materiaal tussen chromosomen plaats.
Historisch gezien wordt dit proces in de literatuur aangeduid met het Angelsaksische woord oversteken. Deze gebeurtenis is een van de resultaten van homologe recombinatie..
De frequentie van oversteken tussen twee genen van hetzelfde chromosoom hangt voornamelijk af van de afstand die tussen hen bestaat; hoe kleiner de fysieke afstand tussen hen, hoe lager de uitwisselingsfrequentie.
Genetisch materiaal wordt continu blootgesteld aan schade, veroorzaakt door endogene en exogene bronnen, zoals straling bijvoorbeeld.
Men schat dat menselijke cellen een aanzienlijk aantal DNA-laesies hebben, in de orde van tientallen tot honderden per dag. Deze laesies moeten worden gerepareerd om mogelijke schadelijke mutaties, replicatie- en transcriptieblokkades en schade op chromosomaal niveau te voorkomen..
Vanuit medisch oogpunt leidt DNA-schade die niet correct wordt hersteld tot de ontwikkeling van tumoren en andere pathologieën.
Homologe recombinatie is een gebeurtenis die DNA-reparatie mogelijk maakt, waardoor verloren sequenties kunnen worden hersteld, waarbij de andere (homologe) DNA-streng als sjabloon wordt gebruikt..
Dit metabolische proces is aanwezig in alle vormen van leven en biedt een betrouwbaar mechanisme dat het mogelijk maakt "gaten" in het DNA, dubbelstrengs breuken en kruisverbindingen tussen DNA-strengen te herstellen..
Een van de meest relevante gevolgen van recombinatie is het genereren van nieuwe genetische variatie. Samen met mutaties zijn het de twee processen die variatie in levende wezens genereren - onthoud dat variatie de grondstof is voor evolutie.
Bovendien biedt het een mechanisme om beschadigde replicatievorken te resetten..
Bij bacteriën zijn er frequente horizontale genoverdrachtsgebeurtenissen. Deze worden geclassificeerd als vervoeging, transformatie en transductie. Hier nemen prokaryoten DNA van een ander organisme en zelfs van verschillende soorten.
Tijdens deze processen vindt homologe recombinatie plaats tussen de ontvangende cel en de donorcel..
Homologe recombinatie begint met de breuk in een van de strengen van het chromosomale DNA-molecuul. Hierna vindt een reeks stappen plaats die worden gekatalyseerd door meerdere enzymen..
Het 3'-uiteinde waar de snede plaatsvindt, wordt binnengedrongen door de homologe dubbele DNA-streng. Het invasieproces is cruciaal. Met "homologe keten" bedoelen we de delen van de chromosomen die dezelfde genen hebben in een lineaire volgorde, hoewel de nucleotidesequenties niet identiek hoeven te zijn.
Deze invasie van de streng plaatst homologe chromosomen tegenover elkaar. Dit fenomeen van het ontmoeten van strengen wordt synaps genoemd (niet te verwarren met de synaps in neuronen, hier wordt de term gebruikt met een andere betekenis).
De synaps impliceert niet noodzakelijkerwijs een direct contact tussen beide homologe sequenties, het DNA kan een tijdje blijven bewegen totdat het het homologe deel vindt. Dit zoekproces wordt homologe uitlijning genoemd..
Dan vindt er een gebeurtenis plaats die "strenginvasie" wordt genoemd. Een chromosoom is een dubbele helix van DNA. Bij homologe recombinatie zoeken twee chromosomen naar hun homologe sequenties. In een van de helices scheiden de strengen zich en deze streng "valt" de dubbele helixstructuur "binnen" en vormt de structuur die de D-lus wordt genoemd..
De D-lus-ketting is verplaatst door de invasie van de breukstreng en paren met de complementaire streng van de originele dubbele helix.
De volgende stap is de vorming van de Holliday-vakbonden. Hier worden de uiteinden van de uitgewisselde strengen met elkaar verbonden. Deze unie heeft het vermogen om in elke richting te bewegen. Het gewricht kan meerdere keren breken en vormen.
Het laatste proces van recombinatie is het oplossen van deze vakbonden en er zijn twee manieren of manieren waarop de cel dit bereikt. Een daarvan is de splitsing van de vereniging of door een proces dat ontbinding wordt genoemd, typisch voor eukaryote organismen..
In het eerste mechanisme regenereert het breken van de Holliday-kruising twee ketens. Bij de andere "ontbinding" -gebeurtenis vindt er een soort ineenstorting plaats in de vakbond..
Een cruciaal eiwit in het recombinatieproces wordt Rad51 genoemd in eukaryote cellen en RecA in Escherichia coli. Het werkt in de verschillende fasen van recombinatie: voor, tijdens en na synaps.
Het Rad51-eiwit vergemakkelijkt de vorming van de fysieke verbinding tussen het binnenvallende DNA en het getemperde DNA. In dit proces wordt het heteroduplex-DNA gegenereerd.
Rad51, en zijn RecA-tegenhanger, katalyseren de zoektocht naar homoloog DNA en de uitwisseling van DNA-strengen. Deze eiwitten hebben het vermogen om samen te binden aan enkelband-DNA..
Er zijn ook paraloge genen (ontstaan uit gen-duplicatiegebeurtenissen in een geslacht van organismen) van Rad51, genaamd Rad55 en Rad57. Bij mensen zijn vijf Rad51 paraloge genen geïdentificeerd, Rad51B, Rad51C, Rad51D, Xrcc2 en Xrcc3..
Omdat recombinatie fysieke binding aan chromosomen vereist, is het een cruciale stap in een goede segregatie tijdens meiose. Als de juiste recombinatie niet plaatsvindt, kan het resultaat een ernstige pathologie zijn.
Niet-disjunctie van chromosomen of fouten in segregatie is een van de meest voorkomende oorzaken van abortussen en anomalieën van chromosomale oorsprong, zoals trisomie van chromosoom 21, die het syndroom van Down veroorzaakt..
Hoewel recombinatie meestal een vrij nauwkeurig proces is, zijn regio's van het genoom die worden herhaald en genen die meerdere kopieën hebben in het genoom, elementen die vatbaar zijn voor een ongelijke crossover.
Deze kruising produceert verschillende klinisch relevante eigenschappen, waaronder veel voorkomende ziekten zoals thalassemie en autisme.
Moleculair biologen hebben geprofiteerd van de kennis van het mechanisme van homologe recombinatie om verschillende technologieën te creëren. Een daarvan maakt de creatie van organismen mogelijk "knock out.
Deze genetisch gemodificeerde organismen maken het mogelijk om de functie van een interessant gen op te helderen.
Een van de methodologieën die worden gebruikt om knockouts Het bestaat uit het onderdrukken van de expressie van het specifieke gen, waarbij het oorspronkelijke gen wordt vervangen door een gemodificeerde of "beschadigde" versie. Het gen wordt uitgewisseld voor de gemuteerde versie door middel van homologe recombinatie.
Naast homologe of legitieme recombinatie zijn er andere soorten uitwisseling van genetisch materiaal.
Wanneer de DNA-gebieden die materiaal uitwisselen niet-allelisch zijn (homologe chromosomen), is het resultaat duplicatie of reductie van genen. Dit proces staat bekend als niet-homologe recombinatie of ongelijke recombinatie..
Gezamenlijk kan genetisch materiaal ook worden uitgewisseld tussen zusterchromatiden op hetzelfde chromosoom. Dit proces vindt plaats in zowel de meiotische als de mitotische deling en wordt ongelijke uitwisseling genoemd..



Niemand heeft nog op dit artikel gereageerd.