
De endoplasmatisch reticulum het is een vliezig cellulair organel dat in alle eukaryote cellen aanwezig is. Dit complexe systeem beslaat ongeveer meer dan de helft van de membranen in een gewone dierencel. De membranen gaan door totdat ze het kernmembraan ontmoeten en een continu element vormen.
Deze structuur is in de vorm van een labyrint door het cytoplasma van de cel verdeeld. Het is een soort netwerk van buisjes die met zakvormige structuren met elkaar zijn verbonden. Binnen het endoplasmatisch reticulum vindt de biosynthese van eiwitten en lipiden plaats. Vrijwel alle eiwitten die naar de buitenkant van de cel moeten worden vervoerd, passeren eerst het reticulum.
Het reticulummembraan is niet alleen verantwoordelijk voor het scheiden van het inwendige van dit organel van de cytoplasmatische ruimte en het bemiddelen van het transport van moleculen tussen deze celcompartimenten; Het is ook betrokken bij de synthese van lipiden, die deel zullen uitmaken van het plasmamembraan van de cel en de membranen van de andere organellen..
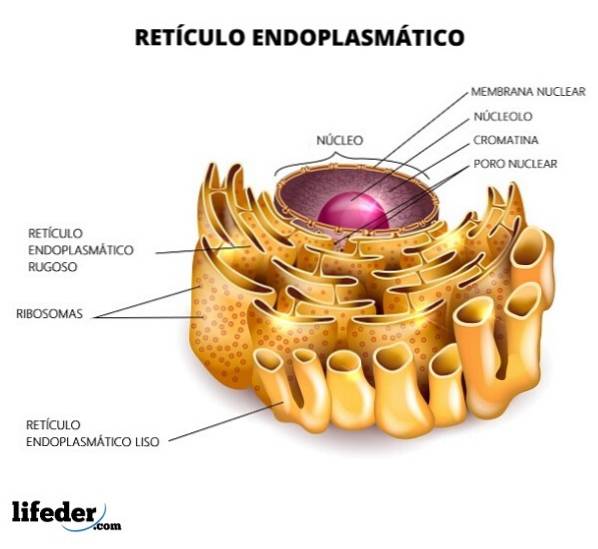
Het reticulum is verdeeld in glad en ruw, afhankelijk van de aanwezigheid of afwezigheid van ribosomen in de membranen. Het ruwe endoplasmatisch reticulum heeft ribosomen die aan het membraan zijn bevestigd (de aanwezigheid van ribosomen geeft het een "ruw" uiterlijk) en de vorm van de tubuli is enigszins recht.
Het gladde endoplasmatisch reticulum van zijn kant mist ribosomen en de vorm van de structuur is veel onregelmatiger. De functie van het ruwe endoplasmatisch reticulum is voornamelijk gericht op de verwerking van eiwitten. Glad is daarentegen verantwoordelijk voor het metabolisme van lipiden.
Artikel index
Het endoplasmatisch reticulum is een vliezig netwerk dat aanwezig is in alle eukaryote cellen. Het is samengesteld uit saccules of stortbakken en buisvormige structuren die een continuüm vormen met het membraan van de kern en zijn verspreid over de cel.
Het reticulum lumen wordt gekenmerkt door hoge concentraties calciumionen, naast een oxiderende omgeving. Beide eigenschappen stellen het in staat zijn functies te vervullen.
Het endoplasmatisch reticulum wordt beschouwd als het grootste organel dat in cellen aanwezig is. Het celvolume van dit compartiment beslaat ongeveer 10% van het celinterieur.
Het ruwe endoplasmatisch reticulum heeft een hoge dichtheid aan ribosomen op het oppervlak. Het is de regio waar alle processen die verband houden met eiwitsynthese en -modificatie plaatsvinden. Het uiterlijk is voornamelijk buisvormig.
Het gladde endoplasmatisch reticulum heeft geen ribosomen. Het is overvloedig aanwezig in celtypen die een actief metabolisme hebben bij de lipidesynthese; bijvoorbeeld in de cellen van de teelballen en eierstokken, die steroïden produceren.
Evenzo wordt het gladde endoplasmatisch reticulum in een vrij hoog aandeel aangetroffen in levercellen (hepatocyten). In dit gebied vindt de productie van lipoproteïnen plaats..
In vergelijking met het ruwe endoplasmatisch reticulum is de structuur gecompliceerder. De overvloed van het gladde versus het ruwe reticulum hangt voornamelijk af van het celtype en zijn functie..
De fysieke architectuur van het endoplasmatisch reticulum is een continu systeem van membranen dat bestaat uit onderling verbonden zakjes en tubuli. Deze membranen strekken zich uit tot in de kern en vormen een enkel lumen.
Het rooster is opgebouwd uit meerdere domeinen. De distributie is geassocieerd met andere organellen, verschillende eiwitten en componenten van het cytoskelet. Deze interacties zijn dynamisch.
Structureel bestaat het endoplasmatisch reticulum uit de nucleaire envelop en het perifere endoplasmatisch reticulum, bestaande uit de tubuli en zakjes. Elke structuur is gerelateerd aan een specifieke functie.
De nucleaire envelop bestaat, net als alle biologische membranen, uit een lipide dubbellaag. Het daardoor afgebakende interieur wordt gedeeld met het perifere reticulum.
De zakjes waaruit het endoplasmatisch reticulum bestaat, zijn plat en vaak gestapeld. Ze bevatten gebogen gebieden aan de randen van de membranen. Het buisvormige netwerk is geen statische entiteit; kan groeien en herstructureren.
Het zak- en tubuli-systeem is aanwezig in alle eukaryote cellen. Het varieert echter in vorm en structuur, afhankelijk van het celtype..
Het netvlies van cellen met belangrijke functies bij de eiwitsynthese bestaat voornamelijk uit zakjes, terwijl de cellen die het meest verband houden met lipidesynthese en calciumsignalering uit een groter aantal tubuli bestaan..
Voorbeelden van cellen met een groot aantal zakjes zijn de secretoire cellen van de pancreas en B-cellen, terwijl spiercellen en levercellen een netwerk van prominente tubuli hebben..
Het endoplasmatisch reticulum is betrokken bij een aantal processen, waaronder eiwitsynthese, handel en vouwing, en modificaties, zoals vorming van disulfidebindingen, glycosylering en de toevoeging van glycolipiden. Bovendien neemt het deel aan de biosynthese van membraanlipiden.
Recente studies hebben het reticulum in verband gebracht met reacties op cellulaire stress en kunnen zelfs apoptoseprocessen induceren, hoewel de mechanismen niet volledig zijn opgehelderd. Al deze processen worden hieronder in detail beschreven:
Het endoplasmatisch reticulum is nauw verbonden met de eiwithandel; specifiek naar eiwitten die naar het buitenland moeten worden gestuurd, naar het Golgi-apparaat, naar lysosomen, naar het plasmamembraan en, logischerwijs, naar die welke tot hetzelfde endoplasmatisch reticulum behoren.
Het endoplasmatisch reticulum is het cellulaire gedrag dat betrokken is bij de synthese van eiwitten die buiten de cel moeten worden gedragen. Deze functie werd in de jaren zestig opgehelderd door een groep onderzoekers die cellen van de alvleesklier bestudeerden die als functie hebben spijsverteringsenzymen af te scheiden..
Deze groep, geleid door George Palade, slaagde erin eiwitten te labelen met radioactieve aminozuren. Op deze manier was het mogelijk om de eiwitten op te sporen en te lokaliseren met een techniek die autoradiografie wordt genoemd..
Radioactief gelabelde eiwitten konden worden herleid tot het endoplasmatisch reticulum. Dit resultaat geeft aan dat het reticulum betrokken is bij de synthese van eiwitten waarvan de uiteindelijke bestemming secretie is..
Later gaan de eiwitten naar het Golgi-apparaat, waar ze worden "verpakt" in blaasjes waarvan de inhoud wordt uitgescheiden..
Het secretieproces vindt plaats omdat het membraan van de blaasjes kan versmelten met het plasmamembraan van de cel (beide zijn lipide van aard). Op deze manier kan de inhoud naar de buitenkant van de cel worden vrijgegeven..
Met andere woorden, uitgescheiden eiwitten (en ook eiwitten die gericht zijn op lysosomen en het plasmamembraan) moeten een specifieke route volgen die het ruwe endoplasmatisch reticulum, het Golgi-apparaat, secretoire blaasjes en tenslotte de buitenkant van de cel omvat..
Eiwitten die bestemd zijn om te worden opgenomen in een bepaald biomembraan (plasmamembraan, Golgi-membraan, lysosoom of reticulum) worden eerst in het reticulummembraan ingebracht en worden niet onmiddellijk in het lumen afgegeven. Ze moeten dezelfde route volgen voor secretoire eiwitten.
Deze eiwitten kunnen door een hydrofobe sector in de membranen worden gelokaliseerd. Dit gebied heeft een reeks van 20 tot 25 hydrobofische aminozuren, die kunnen interageren met de koolstofketens van fosfolipiden. De manier waarop deze eiwitten worden ingebracht, is echter variabel..
Veel eiwitten passeren het membraan maar één keer, terwijl andere dit herhaaldelijk doen. Evenzo kan het in sommige gevallen het einde van de carboxylterminal of de aminoterminal zijn.
De oriëntatie van het eiwit wordt vastgesteld terwijl het peptide groeit en wordt overgebracht naar het endoplasmatisch reticulum. Alle eiwitdomeinen die naar het reticulum lumen wijzen, zullen op hun uiteindelijke locatie aan de buitenkant van de cel worden gevonden..
Moleculen van eiwitachtige aard hebben een driedimensionale conformatie die nodig is om al hun functies uit te voeren..
DNA (deoxyribonucleïnezuur) geeft via een proces dat transcriptie wordt genoemd, zijn informatie door aan een RNA-molecuul (ribonucleïnezuur). Het RNA gaat vervolgens door het translatieproces in de eiwitten. Peptiden worden overgebracht naar het reticulum wanneer het vertaalproces aan de gang is.
Deze aminozuurketens zijn driedimensionaal in het reticulum gerangschikt met behulp van eiwitten die chaperones worden genoemd: een eiwit uit de Hsp70-familie (heat shock-eiwitten of heat shock-eiwitten voor zijn acroniem in het Engels; het getal 70 verwijst naar zijn atoommassa, 70 KDa) genaamd BiP.
Het BiP-eiwit kan binden aan de polypeptideketen en de vouwing ervan mediëren. Evenzo neemt het deel aan de assemblage van de verschillende subeenheden die de quaternaire structuur van eiwitten vormen..
Eiwitten die niet correct zijn gevouwen, worden vastgehouden door het reticulum en blijven gebonden aan BiP of worden afgebroken.
Wanneer de cel wordt blootgesteld aan stressomstandigheden, reageert het reticulum erop en als gevolg daarvan vindt de juiste vouwing van eiwitten niet plaats. De cel kan overgaan op andere systemen en eiwitten produceren die de homeostase van het reticulum in stand houden.
Een disulfidebrug is een covalente binding tussen de sulfhydrylgroepen die deel uitmaken van de aminozuurstructuur cysteïne. Deze interactie is cruciaal voor het functioneren van bepaalde eiwitten; Evenzo definieert het de structuur van de eiwitten waarin ze aanwezig zijn.
Deze bindingen kunnen niet worden gevormd in andere celcompartimenten (bijvoorbeeld in het cytosol), omdat het geen oxiderende omgeving heeft die de vorming ervan bevordert..
Bij de vorming (en het verbreken) van deze bindingen is een enzym betrokken: het eiwit disulfide-isomerase.
In het reticulum vindt het glycosyleringsproces plaats, bij specifieke asparagineresiduen. Net als eiwitvouwing vindt glycosylering plaats terwijl het translatieproces loopt.
De oligosaccharide-eenheden zijn opgebouwd uit veertien suikerresten. Ze worden overgebracht naar asparagine door een enzym genaamd oligosacaryltransferase, dat zich in het membraan bevindt.
Terwijl het eiwit zich in het reticulum bevindt, worden drie glucoseresiduen en één mannoseresidu verwijderd. Deze eiwitten worden naar het Golgi-apparaat gebracht voor verdere verwerking..
Aan de andere kant zijn bepaalde eiwitten niet verankerd aan het plasmamembraan door een deel van hydrofobe peptiden. Daarentegen zijn ze gehecht aan bepaalde glycolipiden die functioneren als een verankeringssysteem en worden ze glycosylfosfatidylinositol genoemd (afgekort als GPI)..
Dit systeem wordt geassembleerd in het reticulummembraan en omvat de binding van de GPI aan de terminale koolstof van het eiwit..
Het endoplasmatisch reticulum speelt een cruciale rol bij de biosynthese van lipiden; specifiek het gladde endoplasmatisch reticulum. Lipiden zijn een onmisbaar onderdeel van de plasmamembranen van cellen.
Lipiden zijn sterk hydrofobe moleculen, dus ze kunnen niet worden gesynthetiseerd in waterige omgevingen. Daarom vindt de synthese ervan plaats in combinatie met reeds bestaande vliezige componenten. Het transport van deze lipiden vindt plaats in blaasjes of door transporteiwitten.
De membranen van eukaryote cellen bestaan uit drie soorten lipiden: fosfolipiden, glycolipiden en cholesterol..
Fosfolipiden zijn afgeleid van glycerol en zijn de belangrijkste structurele bestanddelen. Deze worden gesynthetiseerd in het gebied van het reticulummembraan dat naar het cytosolische gezicht wijst. Verschillende enzymen nemen deel aan het proces.
Het membraan groeit door de integratie van nieuwe lipiden. Dankzij het bestaan van het enzym flipase kan groei optreden in beide helften van het membraan. Dit enzym is verantwoordelijk voor het overbrengen van lipiden van de ene kant van de dubbellaag naar de andere..
De syntheseprocessen van cholesterol en ceramiden komen ook voor in het reticulum. De laatste reist naar het Golgi-apparaat om glycolipiden of sfingomyeline te produceren..
Het calciummolecuul neemt als signaalgever deel aan verschillende processen, of het nu gaat om de fusie of associatie van eiwitten met andere eiwitten of met nucleïnezuren..
Het inwendige van het endoplasmatisch reticulum heeft calciumconcentraties van 100-800 uM. Calciumkanalen en receptoren die calcium afgeven, worden in het reticulum aangetroffen. Calciumafgifte vindt plaats wanneer fosfolipase C wordt gestimuleerd door de activering van G-proteïne-gekoppelde receptoren (GPCR's).
Verder vindt de eliminatie plaats van fosfatidylinositol 4,5-bisfosfaat in diacylglycerol en inositoltrifosfaat; dit laatste is verantwoordelijk voor het vrijkomen van calcium.
Spiercellen hebben een endoplasmatisch reticulum dat gespecialiseerd is in de opslag van calciumionen, het sarcoplasmatisch reticulum genoemd. Het is betrokken bij de processen van spiercontractie en ontspanning.



Niemand heeft nog op dit artikel gereageerd.