
De vetzuren zijn organische macromoleculen afgeleid van koolwaterstoffen, die zijn samengesteld uit lange ketens van koolstof- en waterstofatomen die hydrofoob zijn (ze zijn vetoplosbaar) en die de structurele basis vormen van vetten en lipiden.
Het zijn zeer diverse moleculen die zich van elkaar onderscheiden door de lengte van hun koolwaterstofketens en de aanwezigheid, het aantal, de positie en / of configuratie van hun dubbele bindingen..
In de lipiden van dieren, planten, schimmels en micro-organismen zoals bacteriën en gisten zijn meer dan 100 verschillende klassen vetzuren beschreven en deze worden bij de meeste levende wezens als soort- en weefselspecifiek beschouwd..
De oliën en vetten die de mens dagelijks consumeert zijn van dierlijke of plantaardige oorsprong, ze zijn voornamelijk samengesteld uit vetzuren.
Artikel index

Vetzuurmoleculen vervullen belangrijke functies op cellulair niveau, waardoor ze essentiële componenten zijn en, aangezien sommige niet door dieren kunnen worden gesynthetiseerd, moeten ze ze uit de voeding halen.
Vetzuren zijn ongebruikelijk als vrije soorten in het celcytosol, dus worden ze over het algemeen aangetroffen als onderdeel van andere moleculaire conjugaten zoals:
- Lipiden, in biologische membranen.
- Triglyceriden of vetzuuresters, die dienen als reserves bij planten en dieren.
- Wassen, dit zijn vaste esters van vetzuren met lange ketens en alcoholen.
- Andere soortgelijke stoffen.
Bij dieren worden vetzuren opgeslagen in het cytoplasma van cellen als kleine vetdruppels die zijn samengesteld uit een complex genaamd triacylglycerol, dat niets meer is dan een glycerolmolecuul waaraan het zich in elk van zijn koolstofatomen een vetzuurketen heeft gebonden. door esterkoppelingen.
Hoewel bacteriën korte en meestal enkelvoudig onverzadigde vetzuren bezitten, is het in de natuur gebruikelijk om vetzuren te vinden waarvan de ketens een even aantal koolstofatomen hebben, gewoonlijk tussen 14 en 24, verzadigd, enkelvoudig onverzadigd of meervoudig onverzadigd..
Vetzuren zijn amfipatische moleculen, dat wil zeggen dat ze twee chemisch gedefinieerde gebieden hebben: een hydrofiel polair gebied en een hydrofoob apolair gebied..
Het hydrofobe gebied bestaat uit een lange koolwaterstofketen die, in chemische termen, niet erg reactief is. Het hydrofiele gebied daarentegen bestaat uit een terminale carboxylgroep (-COOH), die zich gedraagt als een zuur.
Deze terminale carboxylgroep of carbonzuur ioniseert in oplossing, is zeer reactief (chemisch gesproken) en is zeer hydrofiel, en vertegenwoordigt dus een covalente bindingsplaats tussen het vetzuur en andere moleculen..
De lengte van de koolwaterstofketens van vetzuren heeft gewoonlijk een even aantal koolstofatomen, en dit hangt nauw samen met het biosynthetische proces waarmee ze worden geproduceerd, aangezien hun groei plaatsvindt in paren koolstofatomen..
De meest voorkomende vetzuren hebben ketens tussen de 16 en 18 koolstofatomen en bij dieren zijn deze ketens niet vertakt.
Vetzuren worden ingedeeld in twee grote groepen volgens de aard van de bindingen waaruit ze bestaan, dat wil zeggen volgens de aanwezigheid van enkele bindingen of dubbele bindingen tussen de koolstofatomen van hun koolwaterstofketens..
Er zijn dus verzadigde en onverzadigde vetzuren.
- Verzadigde vetzuren hebben slechts enkele koolstof-koolstofbindingen en al hun koolstofatomen zijn "verzadigd" of gehecht aan waterstofmoleculen..
- Onverzadigde vetzuren hebben een of meer dubbele koolstof-koolstofbindingen en deze zijn niet allemaal gebonden aan een waterstofatoom.
Onverzadigde vetzuren worden ook verdeeld volgens het aantal onverzadigingen (dubbele bindingen) in enkelvoudig onverzadigde, die met slechts één dubbele binding en meervoudig onverzadigde, die met meer dan één.
Ze hebben meestal tussen de 4 en 26 koolstofatomen die zijn verbonden door enkele bindingen. Het smeltpunt is rechtevenredig met de lengte van de ketting, dat wil zeggen met het molecuulgewicht.
Vetzuren met tussen de 4 en 8 koolstofatomen zijn vloeibaar bij 25 ° C en zijn de vetzuren waaruit eetbare oliën bestaan, terwijl die met meer dan 10 koolstofatomen vast zijn.
Een van de meest voorkomende is laurinezuur, dat overvloedig aanwezig is in palm- en kokosnootolie; palmitinezuur, gevonden in palm, cacao en reuzel, en stearinezuur, gevonden in cacao en gehydrogeneerde oliën.
Het zijn vetzuren met veel meer stabiliteit dan onverzadigde vetzuren, vooral tegen oxidatie, althans onder fysiologische omstandigheden.
Omdat enkele koolstof-koolstofbindingen vrij kunnen roteren, zijn verzadigde vetzuren zeer flexibele moleculen, hoewel sterische hindering de volledig verlengde structuur het meest energetisch stabiel maakt..
Deze vetzuren zijn zeer reactief en vatbaar voor verzadiging en oxidatie. Ze komen veel voor in planten en mariene organismen. Degenen met slechts één dubbele binding staan bekend als enkelvoudig onverzadigd of mono-enoïsch, terwijl degenen met meer dan twee bekend staan als polyenoisch of meervoudig onverzadigd..
De aanwezigheid van dubbele bindingen tussen de koolstofatomen tussen positie 9 en 10 komt veel voor, maar dit betekent niet dat enkelvoudig onverzadigde vetzuren met een onverzadiging op een andere positie niet worden gevonden.
In tegenstelling tot verzadigde vetzuren worden onverzadigde vetzuren niet vermeld van de terminale carboxylgroep, maar volgens de positie van de eerste dubbele C - C-binding. Ze zijn dus verdeeld in twee groepen, omega-6 of ω6-zuren. En omega-3. of ω3.
Omega-6-zuren hebben de eerste dubbele binding op koolstofnummer 6 en omega-3-zuren op koolstofnummer 3. De naam ω wordt gegeven door de dubbele binding die het dichtst bij de laatste methylgroep ligt..
Dubbele bindingen zijn ook te vinden in twee geometrische configuraties die bekend staan als 'cis " Y "trans".
De meeste natuurlijke onverzadigde vetzuren hebben een configuratie "Cis”En de dubbele bindingen van vetzuren die aanwezig zijn in commerciële (gehydrogeneerde) vetten zijn te vinden in "trans".
In meervoudig onverzadigde vetzuren worden twee dubbele bindingen gewoonlijk van elkaar gescheiden door ten minste één methylgroep, dat wil zeggen één koolstofatoom gebonden aan twee waterstofatomen..
Vetzuren hebben meerdere functies in levende organismen en, zoals hierboven vermeld, is een van hun essentiële functies als een essentieel onderdeel van lipiden, de belangrijkste componenten van biologische membranen en een van de drie meest voorkomende biomoleculen in organismen. Levend in combinatie met eiwitten en koolhydraten.
Het zijn ook uitstekende energiesubstraten waardoor grote hoeveelheden energie worden verkregen in de vorm van ATP en andere intermediaire metabolieten..
Aangezien dieren bijvoorbeeld niet in staat zijn koolhydraten op te slaan, vormen vetzuren de belangrijkste bron van energieopslag die afkomstig is van de oxidatie van overmatig verbruikte suikers..
Verzadigde vetzuren met een korte keten in de dikke darm dragen bij aan het stimuleren van de opname van water en natrium-, chloride- en bicarbonaationen; Bovendien hebben ze functies bij de productie van slijm, bij de proliferatie van colonocyten (coloncellen), enz..
Onverzadigde vetzuren komen vooral voor in eetbare plantaardige oliën, die belangrijk zijn in de voeding van alle mensen.

Anderen nemen deel als liganden van sommige eiwitten met enzymatische activiteiten, dus ze zijn belangrijk met betrekking tot hun effecten op het energiemetabolisme van de cellen waarin ze worden aangetroffen..
De afbraak van vetzuren staat bekend als β-oxidatie en vindt plaats in de mitochondriën van eukaryote cellen. Biosynthese daarentegen vindt plaats in het cytosol van dierlijke cellen en in de chloroplasten (fotosynthetische organellen) van plantencellen..
Het is een proces dat afhankelijk is van acetyl-CoA, malonyl-CoA en NADPH, het komt voor in alle levende organismen en bij "hogere" dieren zoals zoogdieren. Het is bijvoorbeeld erg belangrijk in lever- en vetweefsel, maar ook in de borstklieren..
De NADPH die voor deze route wordt gebruikt, is voornamelijk het product van de NADP-afhankelijke oxidatiereacties van de pentosefosfaatroute, terwijl acetyl-CoA uit verschillende bronnen kan komen, bijvoorbeeld van de oxidatieve decarboxylering van pyruvaat, van de Krebs-cyclus en β-oxidatie. van vetzuren.
De biosyntheseroute wordt, net als die van β-oxidatie, in alle cellen sterk gereguleerd door allostere effectoren en covalente modificaties van enzymen die deelnemen aan de regulatie..
De route begint met de vorming van een metabolisch tussenproduct dat bekend staat als malonyl-CoA uit een acetyl-CoA-molecuul en wordt gekatalyseerd door een multifunctioneel enzym genaamd acetyl-CoA-carboxylase..
Deze reactie is een additiereactie van een biotine-afhankelijk carboxylmolecuul (-COOH, carboxylering) en verloopt in twee stappen:
Bij dieren vindt de vorming van koolhydraatketens van vetzuren verder plaats door opeenvolgende condensatiereacties die worden gekatalyseerd door een multimeer en multifunctioneel enzym dat bekend staat als vetzuursynthase..
Dit enzym katalyseert de condensatie van een acetyl-CoA-eenheid en meerdere malonyl-CoA-moleculen die worden geproduceerd uit de acetyl-CoA-carboxylasereactie, een proces waarbij één molecuul CO2 vrijkomt voor elke malonyl-CoA die het toevoegt.
De groeiende vetzuren worden veresterd tot een eiwit genaamd "acyl carrier protein" of ACP, dat thioesters vormt met acylgroepen. Aan E coli dit eiwit is een polypeptide van 10 kDa, maar bij dieren maakt het deel uit van het vetzuursynthasecomplex.
Bij het verbreken van deze thioesterbindingen komen grote hoeveelheden energie vrij, waardoor thermodynamisch gesproken condensatiestappen in de biosynthetische route kunnen optreden..
In bacteriën komt de vetzuursynthase-activiteit eigenlijk overeen met zes onafhankelijke enzymen die acetyl-coA en malonyl-coA gebruiken om vetzuren te vormen en waarmee zes verschillende enzymatische activiteiten zijn geassocieerd..
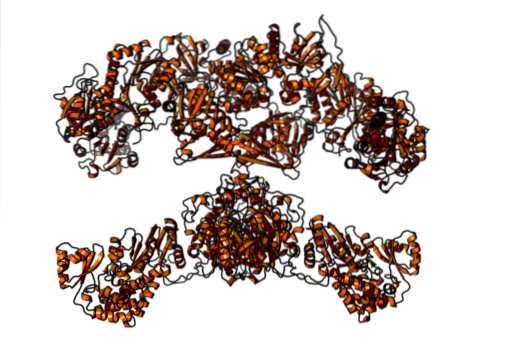
Bij zoogdieren daarentegen is vetzuursynthase een multifunctioneel homodimeer enzymcomplex met een molecuulgewicht van ongeveer 500 kDa, dat zes verschillende katalytische activiteiten heeft en waarmee het acyldragereiwit zich associeert..
De thiolgroepen in de cysteïneresiduen die verantwoordelijk zijn voor de binding van metabolische tussenproducten aan het ACP-enzym, moeten vóór het begin van de synthese worden beladen met de nodige acylgroepen..
Hiervoor wordt de acetylgroep van acetyl-coA overgebracht naar de thiolgroep (-SH) van een van de cysteïneresten van de ACP-subeenheid van vetzuursynthase. Deze reactie wordt gekatalyseerd door de ACP-acyltransferase-subeenheid..
De acetylgroep wordt vervolgens overgebracht van het ACP naar een ander cysteïneresidu op de katalytische plaats van een andere enzymsubeenheid van het complex dat bekend staat als β-ketoacyl-ACP-synthase. Het enzymcomplex wordt dus "geprimed" om met synthese te beginnen..
Malonyl-CoA dat wordt geproduceerd door acetyl-CoA-carboxylase wordt overgebracht naar de thiolgroep in ACP en tijdens deze reactie gaat het CoA-gedeelte verloren. De reactie wordt gekatalyseerd door de malonyl-ACP-transferase-subeenheid van het vetzuursynthasecomplex, dat vervolgens malonyl-ACP produceert.
Tijdens dit proces wordt de malonylgroep gekoppeld aan ACP en β-ketoacyl-ACP-synthase via respectievelijk een ester en een andere sulfhydrylbinding..
Het enzym β-ketoacyl-ACP-synthase katalyseert de overdracht van de acetylgroep die eraan was gehecht in de "priming" -stap naar de 2-koolstofatomen van de malonylgroep die, in de vorige stap, werd overgedragen aan de ACP..
Tijdens deze reactie komt een CO2-molecuul vrij uit malonyl, wat overeenkomt met het CO2 dat wordt geleverd door bicarbonaat bij de carboxyleringsreactie van acetyl-CoA-carboxylase. Acetoacetyl-ACP wordt vervolgens geproduceerd.
De β-ketoacyl-ACP-reductase-subeenheid katalyseert de NADPH-afhankelijke reductie van acetoacetyl-ACP, waardoor D-β-hydroxybutyryl-ACP wordt gevormd.
In deze stap wordt trans-α, β-acyl-ACP of ∆2-onverzadigde-acyl-ACP (cratonyl-ACP) gevormd, een product van de dehydratie van D-β-hydroxybutyryl-ACP door de werking van de enoyl- subeenheid ACP-hydratase.
Later wordt cratonyl-ACP gereduceerd tot butyryl-ACP door een NADPH-afhankelijke reactie die wordt gekatalyseerd door de enoyl-ACP-reductase-subeenheid. Deze reactie voltooit de eerste van zeven cycli die nodig zijn om palmitoyl-ACP te produceren, een voorloper van bijna alle vetzuren..
De butyrylgroep wordt overgebracht van de ACP naar de thiolgroep van een cysteïneresidu in β-ketoacyl-ACP-synthase, waarmee de ACP een andere malonylgroep van malonyl-CoA kan accepteren.
Op deze manier is de reactie die optreedt de condensatie van malonyl-ACP met buturyl-β-ketoacyl-ACP-synthase, wat aanleiding geeft tot β-ketohexanoyl-ACP + CO2.
Het palmitoyl-ACP dat ontstaat uit de volgende stappen (na toevoeging van 5 extra malonyl-eenheden) kan worden vrijgemaakt als vrij palmitinezuur dankzij de activiteit van het thioesterase-enzym, het kan worden overgebracht naar CoA of worden opgenomen in fosfatidinezuur voor het fosfolipide. en triacylglyceridesyntheseroute.
De vetzuursynthase van de meeste organismen stopt bij de synthese van palmitoyl-ACP, aangezien de katalytische plaats van de β-ketoacyl-ACP-synthase-subeenheid een configuratie heeft waarin alleen vetzuren van die lengte kunnen worden ondergebracht..
Deze komen relatief vaak voor in mariene organismen en worden ook gesynthetiseerd door een vetzuursynthasecomplex. De "priming" -reactie vindt echter plaats met een langer molecuul, propionyl-ACP, met drie koolstofatomen..
Palmitinezuur dient, zoals besproken, als een precursor voor veel verzadigde en onverzadigde vetzuren met een langere keten. Het proces van "verlenging" van vetzuren vindt plaats in de mitochondriën, terwijl de introductie van onverzadigingen hoofdzakelijk plaatsvindt in het endoplasmatisch reticulum..
Veel organismen zetten hun verzadigde vetzuren om in onverzadigde vetzuren als aanpassing aan lage omgevingstemperaturen, omdat ze hierdoor het smeltpunt van lipiden onder kamertemperatuur kunnen houden..
Veel van de eigenschappen van vetzuren zijn afhankelijk van hun ketenlengte en de aanwezigheid en het aantal onverzadigingen:
- Onverzadigde vetzuren hebben een lager smeltpunt dan verzadigde vetzuren van dezelfde lengte.
- De lengte van vetzuren (het aantal koolstofatomen) is omgekeerd evenredig met de vloeibaarheid of flexibiliteit van het molecuul, dat wil zeggen dat de "kortere" moleculen vloeibaarder zijn en vice versa..
Over het algemeen zijn vloeibare vetstoffen samengesteld uit vetzuren met een korte keten met de aanwezigheid van onverzadigingen.
Planten hebben overvloedige hoeveelheden onverzadigde vetzuren, evenals dieren die bij zeer lage temperaturen leven, omdat deze, als componenten van de lipiden die aanwezig zijn in celmembranen, ze onder deze omstandigheden een grotere vloeibaarheid geven..
Onder fysiologische omstandigheden veroorzaakt de aanwezigheid van een dubbele binding in de koolwaterstofketen van een vetzuur een kromming van ongeveer 30 °, waardoor deze moleculen een grotere ruimte innemen en de sterkte van hun van der Waals-interacties verminderen..
De aanwezigheid van dubbele bindingen in vetzuren geassocieerd met lipidemoleculen heeft directe effecten op de mate van "verpakking" die deze kunnen hebben in de membranen waartoe ze behoren en hebben dus ook effecten op membraaneiwitten..
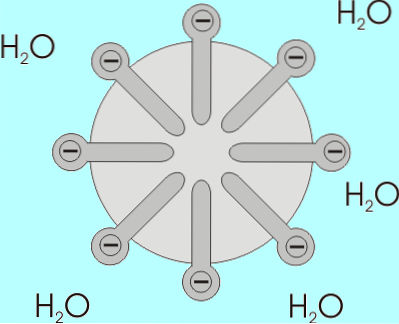
De oplosbaarheid van vetzuren neemt af naarmate hun ketenlengte toeneemt, dus ze zijn omgekeerd evenredig. In waterige en lipidenmengsels associëren vetzuren zich in structuren die bekend staan als micellen..
Een micel is een structuur waarin de alifatische ketens van vetzuren zijn "ingesloten", waardoor alle watermoleculen worden "verdreven" en op het oppervlak waarvan de carboxylgroepen zich bevinden..
De nomenclatuur van vetzuren kan enigszins complex zijn, vooral als men verwijst naar de gewone namen die ze krijgen, die vaak verband houden met een fysisch-chemische eigenschap, met de plaats waar ze worden gevonden of andere kenmerken.
Veel auteurs zijn van mening dat, aangezien deze moleculen zijn geïoniseerd bij fysiologische pH dankzij de terminale carboxylgroep, men naar hen moet verwijzen als "carboxylaten" met gebruikmaking van de terminatie ".ato ".
Volgens het IUPAC-systeem wordt de opsomming van de koolstofatomen van een vetzuur gemaakt van de carboxylgroep aan het polaire uiteinde van het molecuul en worden de eerste twee koolstofatomen die aan deze groep zijn bevestigd, respectievelijk α en β genoemd. De terminale methyl van de keten bevat het koolstofatoom ω.
Over het algemeen krijgen ze in de systematische nomenclatuur de naam van de "moederkoolwaterstof" (de koolwaterstof met hetzelfde aantal koolstofatomen) en het einde "of" voor "Oico", als het een onverzadigd vetzuur is, wordt het einde toegevoegd "Enoic".
Beschouw bijvoorbeeld het geval van een C18 (C18) vetzuur:
- Omdat de koolwaterstof met hetzelfde aantal koolstofatomen bekend staat als octadecaan, wordt het verzadigde zuur "octadecaanzuur" O goed "octadecanoaat”En de algemene naam is stearinezuur.
- Als je een dubbele binding hebt tussen een paar koolstofatomen in je structuur, staat deze bekend als 'octadeceenzuur
- Als het twee dubbele bindingen c - c heeft, wordt het aangeroepen "Octadecadienoic acid" en als je er drie hebt "octadecatrieenzuur.
Als je de nomenclatuur wilt samenvatten, dan wordt 18: 0 gebruikt voor het 18-koolstofvetzuur en geen dubbele bindingen (verzadigd) en, afhankelijk van de mate van onverzadiging, dan wordt in plaats van nul 18: 1 geschreven voor een molecuul met een onverzadiging, 18: 2 voor een met twee onverzadigingen enzovoort.
Als je wilt specificeren tussen welke koolstofatomen de dubbele bindingen in onverzadigde vetzuren zijn, gebruik dan het symbool ∆ met een numeriek superscript dat de plaats van onverzadiging aangeeft en het voorvoegsel "Cis"Of "trans", afhankelijk van de configuratie hiervan.



Niemand heeft nog op dit artikel gereageerd.