De tertiaire structuur van eiwitten Het is de driedimensionale conformatie die polypeptideketens verwerven wanneer ze zichzelf vouwen. Deze conformatie verschijnt door interacties tussen de zijketens van de aminozuurresiduen van het polypeptide. Zijketens kunnen interageren ongeacht hun positie in het eiwit.
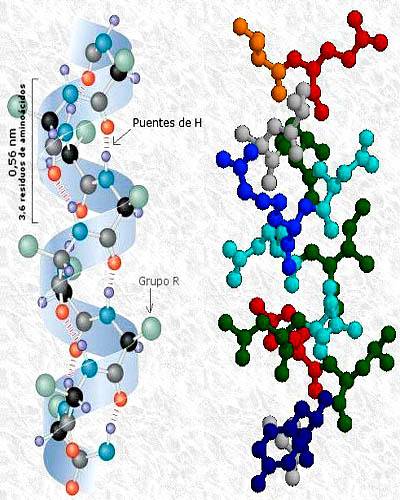
Omdat het afhangt van de interacties tussen de R-groepen, vertoont de tertiaire structuur niet-repetitieve aspecten van de keten, aangezien deze groepen verschillend zijn voor elk aminozuurresidu. De secundaire structuur is daarentegen afhankelijk van de carboxyl- en aminogroepen, die in alle aminozuren aanwezig zijn.
Sommige auteurs suggereren dat fibreuze eiwitten een eenvoudige tertiaire structuur hebben, maar toch wijzen andere auteurs erop dat deze structuur typerend is voor globulaire eiwitten..
Artikel index
In vezelachtige eiwitten zijn de polypeptideketens gerangschikt in de vorm van lange filamenten of lange vellen; ze bestaan over het algemeen uit een enkel type secundaire structuur. Deze secundaire structuur is in de meeste gevallen belangrijker dan de tertiaire structuur bij het bepalen van de vorm van het eiwit..
De biologische functie is structureel en geeft kracht en / of elasticiteit aan de organen en structuren waar ze worden aangetroffen, terwijl ze bij elkaar blijven. Alle vezelige eiwitten zijn onoplosbaar in water, vanwege de grote hoeveelheid hydrofobe aminozuurresten die ze bevatten.
Deze vezelige eiwitten omvatten keratine en collageen. De eerste worden aangetroffen in bindweefsels en in structuren zoals haren, nagels (α-keratine), schubben en veren (β-keratine). Collageen wordt op zijn beurt onder andere aangetroffen in botten, pezen en huid.
Deze eiwitten maken deel uit van de zogenaamde intermediaire filamenteiwitten, die een belangrijke rol spelen in het cytoskelet van meercellige organismen. Bovendien zijn ze het hoofdbestanddeel van haar, nagels, wol, hoorns, hoeven en een van de belangrijkste eiwitten van dierenhuid..
De structuur van het molecuul is een α-helix. Twee α-keratinestrengen kunnen parallel worden gerangschikt en over elkaar worden gewikkeld, waarbij hun hydrofobe R-groepen met elkaar in wisselwerking staan. Op deze manier ontstaat een superhelische structuur of bal met een linksdraaiende wikkeling..
De tertiaire structuur van α-keratine is eenvoudig en wordt gedomineerd door de secundaire structuur van α-helix. Aan de andere kant is de quaternaire structuur ook aanwezig, aangezien twee moleculen deelnemen aan de superhelicale structuur, die interageren via niet-covalente bindingen..
De primaire structuur is vergelijkbaar met die van α-keratines, maar hun secundaire structuur wordt gedomineerd door β-platen. Ze zijn het hoofdbestanddeel van reptielenschubben en vogelveren..
Dit eiwit kan meer dan 30% van de totale eiwitmassa van sommige dieren vertegenwoordigen. Gevonden in kraakbeen, botten, pezen, het hoornvlies en de huid, onder andere weefsels.
De secundaire structuur van collageen is uniek en wordt weergegeven door een linkshandige helix met 3,3 aminozuurresiduen voor elke draai. Drie linkshandige helixketens (α-ketens) wikkelen zich om elkaar heen, waardoor een rechtshandige supercoiled molecuul ontstaat, door sommige auteurs bekend als tropocollageen..
Tropocollageenmoleculen komen samen om een collageenvezel te vormen die een hoge sterkte heeft, superieur is aan die van staal en vergelijkbaar is met die van hoogwaardig koper.
Andere soorten vezelige eiwitten zijn fibroïne en elastine. De eerste bestaat uit β-vellen, voornamelijk bestaande uit glycine, alanine en serine..
De zijketens van deze aminozuren zijn klein van formaat, dus ze kunnen stevig worden verpakt. Het resultaat is een vezel die zowel zeer resistent als nauwelijks rekbaar is..
In elastine vervangt valine op zijn beurt serine van de belangrijkste samenstellende aminozuren. In tegenstelling tot fibroïne is elastine zeer rekbaar, vandaar de naam. In de samenstelling van het molecuul werkt lysine ook, dat kan deelnemen aan verknopingen waardoor elastine zijn vorm terugkrijgt wanneer de spanning ophoudt.
Bolvormige eiwitten zijn, in tegenstelling tot vezelachtige, oplosbaar en hebben over het algemeen verschillende soorten secundaire structuren. Hierin zijn echter de driedimensionale conformaties die ze verwerven wanneer ze op zichzelf vouwen belangrijker (tertiaire structuur).
Deze specifieke driedimensionale conformaties verlenen aan elk eiwit een specifieke biologische activiteit. De belangrijkste functie van deze eiwitten is regulerend, zoals gebeurt bij enzymen.
De tertiaire structuur van globulaire eiwitten heeft enkele belangrijke kenmerken:
- Globulaire eiwitten zijn compact dankzij de verpakking door de polypeptideketen te vouwen.
- De verre aminozuurresiduen in de primaire structuur van de polypeptideketens blijven dicht bij elkaar en kunnen met elkaar communiceren vanwege de vouwing.
- Grotere globulaire eiwitten (meer dan 200 aminozuren) kunnen verschillende compacte segmenten hebben, onafhankelijk van elkaar en met specifieke functies, en elk van deze segmenten wordt een domein genoemd. Een domein kan tussen de 50 en 350 aminozuurresiduen bevatten.
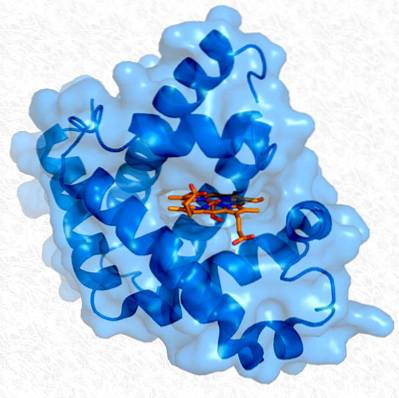
Zoals reeds opgemerkt, vertonen eiwitten bepaalde vormen van vouwing, waardoor ze ook bijzondere kenmerken krijgen. Dit vouwen is niet willekeurig en wordt begunstigd door zowel de primaire en secundaire structuur als door enkele niet-covalente interacties, en er zijn ook enkele fysieke beperkingen aan het vouwen, waarvoor enkele regels zijn opgesteld:
- Alle bolvormige proteïnen hebben gedefinieerde distributiepatronen, met de hydrofobe R-groepen gericht naar het inwendige van het molecuul en de hydrofiele residuen in de buitenste laag. Dit vereist ten minste twee lagen secundaire structuur. De β-α-β-lus en het α-α-hoekpunt kunnen deze twee lagen bieden.
- De β-vellen zijn over het algemeen gerangschikt in een linkshandige opgerolde vorm..
- In een polypeptideketen kunnen verschillende wendingen optreden die van de ene secundaire structuur naar de andere gaan, zoals β- of γ-wendingen, die de richting van de keten met vier aminozuurresiduen of minder kunnen omkeren..
- Bolvormige eiwitten bezitten α-helices, β-vellen, bochten en onregelmatig gestructureerde segmenten.
Als een eiwit zijn natuurlijke (natuurlijke) driedimensionale structuur verliest, verliest het zijn biologische activiteit en de meeste van zijn specifieke eigenschappen. Dit proces staat bekend onder de naam denaturatie.
Denaturatie kan optreden wanneer natuurlijke omgevingsomstandigheden veranderen, bijvoorbeeld door temperatuur of pH te variëren. Bij veel eiwitten is het proces onomkeerbaar; andere kunnen echter spontaan hun natuurlijke structuur terugkrijgen wanneer de normale omgevingsomstandigheden worden hersteld..



Niemand heeft nog op dit artikel gereageerd.