De tussenliggende filamenten, ook bekend in de literatuur als "IF's" (uit het Engels Tussenliggende filamenten), zijn een familie van onoplosbare cytosolische fibreuze eiwitten die aanwezig zijn in alle cellen van meercellige eukaryoten.
Ze maken deel uit van het cytoskelet, een intracellulair filamenteus netwerk dat voornamelijk verantwoordelijk is voor de ondersteuning van de celstructuur en voor verschillende metabolische en fysiologische processen zoals transport van blaasjes, celbeweging en verplaatsing, enz..
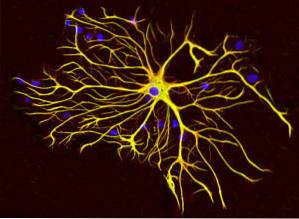
Samen met microtubuli en microfilamenten nemen intermediaire filamenten deel aan de ruimtelijke organisatie van intracellulaire organellen, aan de processen van endocytose en exocytose, en ook aan de processen van celdeling en intercellulaire communicatie..
De eerste intermediaire filamenten die werden bestudeerd en beschreven, waren keratine, een van de eerste soorten eiwitten waarvan de structuur in de jaren dertig door röntgendiffractie werd geanalyseerd..
Het concept van tussenliggende filamenten werd echter in de jaren tachtig geïntroduceerd door Lazarides, die ze beschreef als complexe "mechanische integratoren van celruimte", gekenmerkt door hun onoplosbaarheid en hun vermogen om weer in elkaar te zetten. in vitro na denaturatie.
Ze worden door veel auteurs beschouwd als stress-"buffer" -elementen voor dierlijke cellen, aangezien ze flexibeler filamenten zijn dan microtubuli en microfilamenten. Ze worden niet alleen in het cytoskelet aangetroffen, maar maken ook deel uit van het nucleoskelet.
In tegenstelling tot de andere vezelcomponenten van het cytoskelet, nemen de tussenliggende filamenten niet rechtstreeks deel aan de processen van celmobiliteit, maar functioneren ze eerder in het structurele onderhoud en de mechanische weerstand van cellen..
Artikel index
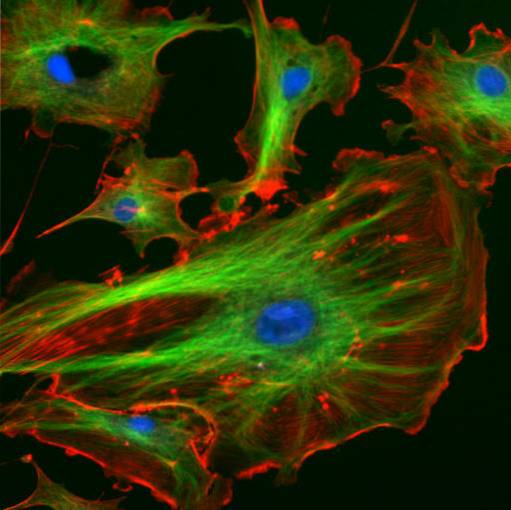
De tussenliggende filamenten hebben een diameter van ongeveer 10 nm, een structureel kenmerk waarvoor ze zijn genoemd, aangezien hun grootte ligt tussen de afmetingen die overeenkomen met myosine- en actine-filamenten, die respectievelijk tussen 25 en 7 nm liggen..
Ze verschillen structureel van de andere twee typen cytoskeletfilamenten, die bolvormige eiwitpolymeren zijn, doordat hun samenstellende eenheden verschillende lange α-helix-vezelige eiwitten zijn die samen clusteren om touwachtige structuren te vormen..
Alle eiwitten die de tussenliggende filamenten vormen, hebben een vergelijkbare moleculaire organisatie, bestaande uit een α-helix- of "touw" -domein met verschillende hoeveelheden "spiraalvormende" segmenten van dezelfde grootte..
Dit helixvormige domein wordt geflankeerd door een N-terminale niet-helixvormige "kop" en een niet-helixvormige "staart" aan het C-terminale uiteinde, die beide variëren in zowel grootte als aminozuursequentie..
Binnen de opeenvolging van deze twee uiteinden bevinden zich de consensusmotieven die gemeenschappelijk zijn voor de 6 bekende typen intermediaire filamenten.
Bij gewervelde dieren is het "akkoord" -domein van cytosolische intermediaire filament-eiwitten ongeveer 310 aminozuurresiduen, terwijl cytosolische eiwitten van ongewervelde en nucleaire lamina ongeveer 350 aminozuren lang zijn..
Tussenliggende filamenten zijn "zelfassemblerende" structuren die geen enzymatische activiteit bezitten, waardoor ze zich ook onderscheiden van hun tegenhangers in het cytoskelet (microtubuli en microfilamenten)..
Deze structuren worden aanvankelijk samengesteld als tetrameren van de filamenteuze eiwitten waaruit ze bestaan, uitsluitend onder invloed van eenwaardige kationen..
Deze tetrameren zijn 62 nm lang en hun monomeren associëren zich lateraal met elkaar om "lengte-eenheden" (UFL) te vormen. filamenten van eenheidslengte), die bekend staat als fase 1 van de montage, die zeer snel verloopt.
UFL's zijn de voorlopers van lange filamenten en, aangezien de dimeren waaruit ze bestaan op een antiparallelle en verspringende manier met elkaar zijn verbonden, hebben deze eenheden een centraal domein met twee flankerende domeinen waardoor fase 2 van verlenging plaatsvindt., Waar de longitudinale vereniging van andere UFL's komen voor.
Tijdens wat fase 3 van de assemblage wordt genoemd, vindt radiale verdichting van de diameter van de filamenten plaats, wat rijpe tussenliggende filamenten produceert met een diameter van ongeveer 10 nm..
De functies van de tussenliggende filamenten hangen sterk af van het beschouwde celtype en, in het geval van dieren (inclusief mensen), wordt hun expressie op een weefselspecifieke manier gereguleerd, daarom hangt het ook af van het type weefsel..
Epithelia, spieren, mesenchymale en gliacellen en neuronen hebben verschillende soorten filamenten, gespecialiseerd volgens de functie van de cellen waartoe ze behoren..
Van deze functies zijn de belangrijkste het structurele onderhoud van de cellen en de weerstand tegen verschillende mechanische spanningen, aangezien deze structuren een zekere elasticiteit hebben waardoor ze verschillende soorten krachten kunnen opvangen die op de cellen worden uitgeoefend..
De eiwitten die de intermediaire filamenten vormen, behoren tot een grote en heterogene familie van filamenteuze eiwitten die chemisch verschillend zijn, maar die worden onderscheiden in zes klassen op basis van hun sequentiehomologie (I, II, III, IV, V en VI).
Hoewel het niet erg gebruikelijk is, kunnen verschillende soorten cellen, onder zeer specifieke omstandigheden (ontwikkeling, celtransformatie, groei, enz.), Meer dan één klasse van intermediaire filamentvormende eiwitten tot expressie brengen.
Keratines zijn verantwoordelijk voor de meeste eiwitten in de tussenliggende filamenten en bij mensen zijn ze goed voor meer dan driekwart van de tussenliggende filamenten..
Ze hebben molecuulgewichten die variëren tussen 40 en 70 kDa en verschillen van andere intermediaire filamenteiwitten door hun hoge gehalte aan glycine- en serineresiduen..
Ze staan bekend als zure en basische keratine vanwege hun iso-elektrische punten, die tussen 4,9 en 5,4 liggen voor zure keratine en tussen 6,1 en 7,8 voor basische keratine..
In deze twee klassen zijn ongeveer 30 eiwitten beschreven en deze zijn vooral aanwezig in epitheelcellen, waar beide typen eiwitten "co-polymeriseren" en samengestelde filamenten vormen..
Veel van de keratines van het tussenliggende filamentgeval I worden aangetroffen in structuren zoals haar, nagels, hoorns, pennen en klauwen, terwijl die van klasse II het meest voorkomen in het cytosol..
Desmine is een zuur eiwit van 53 kDa dat, afhankelijk van de mate van fosforylering, verschillende varianten heeft.
Sommige auteurs hebben desmine-filamenten ook "intermediaire spierfilamenten" genoemd, aangezien hun aanwezigheid vrij beperkt is, hoewel in kleine hoeveelheden, tot alle soorten spiercellen..
In myofibrillen wordt desmine aangetroffen in de Z-lijn, en daarom wordt aangenomen dat dit eiwit bijdraagt aan de contractiele functies van spiervezels door te functioneren op de kruising van myofibrillen en het plasmamembraan..
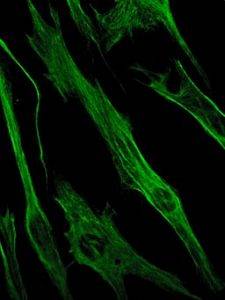
Vimentine is op zijn beurt een eiwit dat aanwezig is in mesenchymale cellen. De tussenliggende filamenten die door dit eiwit worden gevormd, zijn flexibel en blijken bestand te zijn tegen veel van de conformatieveranderingen die optreden tijdens de celcyclus..
Het wordt aangetroffen in fibroblasten, gladde spiercellen, witte bloedcellen en andere cellen van de bloedsomloop van dieren..
Deze klasse van intermediaire filamenten, ook bekend als "neurofilamenten", omvat een van de fundamentele structurele elementen van neuronale axonen en dendrieten; worden vaak geassocieerd met de microtubuli waaruit deze structuren ook bestaan.
De neurofilamenten van gewervelde dieren zijn geïsoleerd, wat aangeeft dat het een triplet is van eiwitten van 200, 150 en 68 kDa die deelnemen aan de assemblage in vitro.
Ze verschillen van andere tussenliggende filamenten doordat ze zijarmen hebben als "aanhangsels" die uitsteken vanaf de omtrek ervan en die functioneren in de interactie tussen naburige filamenten en andere structuren..
Gliacellen produceren een speciaal type tussenfilamenten die bekend staan als gliale tussenfilamenten, die structureel verschillen van neurofilamenten doordat ze zijn samengesteld uit een enkel eiwit van 51 kDa en verschillende fysisch-chemische eigenschappen hebben..
Alle laminae die deel uitmaken van het nucleoskelet zijn eigenlijk intermediaire filamenteiwitten. Deze hebben een molecuulgewicht tussen 60 en 75 kDa en worden aangetroffen in de kernen van alle eukaryote cellen.
Ze zijn essentieel voor de interne organisatie van de nucleaire regio's en voor veel van de functies van dit organel die essentieel zijn voor het bestaan van eukaryoten..
Dit type tussenfilament weegt ongeveer 200 kDa en wordt voornamelijk aangetroffen in stamcellen van het centrale zenuwstelsel. Ze komen tot uiting tijdens neurale ontwikkeling.
Er zijn meerdere ziekten bij mensen die verband houden met tussenliggende filamenten.
Bij sommige soorten kanker, zoals kwaadaardige melanomen of borstcarcinomen, leidt de gezamenlijke expressie van tussenliggende filamenten van vimentine en keratine tot de differentiatie of onderlinge omzetting van epitheliale en mesenchymale cellen..
Experimenteel is aangetoond dat dit fenomeen de migrerende en invasieve activiteit van kankercellen verhoogt, wat belangrijke implicaties heeft voor de metastatische processen die kenmerkend zijn voor deze aandoening..
Eriksson et al. (2009) geven een overzicht van de verschillende soorten ziekten en hun relatie met specifieke mutaties in de genen die betrokken zijn bij de vorming van de zes soorten intermediaire filamenten..
Ziekten die verband houden met mutaties in de genen die coderen voor de twee soorten keratine zijn epidermolysis bullosa, epidermolytische hyperkeratose, corneadystrofie, keratodermie en vele andere..
Type III intermediaire filamenten zijn betrokken bij talrijke cardiomyopathieën en bij verschillende spierziekten die voornamelijk verband houden met dystrofieën. Bovendien zijn ze ook verantwoordelijk voor dominante cataracten en sommige soorten sclerose.
Veel neurologische syndromen en aandoeningen zijn geassocieerd met type IV-filamenten, zoals Parkinson. Evenzo zijn genetische defecten in type V- en VI-filamenten verantwoordelijk voor de ontwikkeling van verschillende autosomale ziekten en houden ze verband met het functioneren van de celkern..
Voorbeelden hiervan zijn onder meer het Hutchinson-Gilford-progeria-syndroom, Emery-Dreifuss-spierdystrofie..


Niemand heeft nog op dit artikel gereageerd.