Met "niet-Mendeliaanse overerving'We bedoelen elk patroon van overerving waarin overgeërfde karakters niet gescheiden zijn in overeenstemming met de bepalingen van de wetten van Mendel..
In 1865 voerde Gregor Mendel, die beschouwd werd als de "vader van de genetica", een reeks experimentele kruisingen uit met erwtenplanten, waarvan de resultaten hem ertoe brachten enkele postulaten (de wetten van Mendel) voor te stellen die een logische verklaring probeerden te geven voor overerving. de karakters tussen ouders en kinderen.
Deze scherpzinnige Oostenrijkse monnik observeerde zorgvuldig de segregatie van oudergenen en hun verschijning in de nakomelingen als dominante en recessieve karakters. Bovendien bepaalde hij de wiskundige patronen die de overerving van de ene generatie op de andere beschreven en deze bevindingen werden "geordend" in de vorm van 3 fundamentele wetten:
- De wet van dominantie
- De wet van de scheiding van karakters en
- De wet van onafhankelijke distributie.
De successen en conclusies van Mendel bleven jarenlang verborgen, tot hun herontdekking aan het begin van de 20e eeuw..

In die tijd hield de wetenschappelijke gemeenschap echter een ietwat sceptische positie in met betrekking tot deze wetten, aangezien ze de erfelijkheidspatronen in geen enkele dier- of plantensoort leken te verklaren, vooral niet in die kenmerken die door meer dan één locus werden bepaald..
Hierdoor classificeerden de eerste genetici de erfelijke patronen die werden waargenomen als 'Mendeliaans' (die konden worden verklaard door de segregatie van eenvoudige, dominante of recessieve allelen die tot dezelfde locus behoren) en 'niet-Mendeliaans' (die niet-Mendeliaans). kon zo gemakkelijk worden uitgelegd).
Artikel index
Mendeliaanse overerving verwijst naar een erfelijk patroon dat voldoet aan de wetten van segregatie en onafhankelijke distributie, volgens welke een gen dat wordt geërfd van een van de ouders uitscheidt in gameten met een equivalente frequentie of, liever, met dezelfde waarschijnlijkheid..
De belangrijkste Mendeliaanse overervingspatronen die zijn beschreven voor sommige ziekten zijn: autosomaal recessief, autosomaal dominant en gekoppeld aan het X-chromosoom, die worden toegevoegd aan de dominantie- en recessiviteitspatronen beschreven door Mendel.
Deze werden echter gepostuleerd met betrekking tot zichtbare eigenschappen en niet met betrekking tot genen (er moet worden opgemerkt dat sommige allelen coderen voor eigenschappen die segregeren als dominant, terwijl andere dezelfde eigenschappen kunnen coderen, maar deze worden gescheiden als recessieve genen).
Uit het bovenstaande volgt dat niet-Mendeliaanse overerving eenvoudigweg bestaat uit elk erfelijk patroon dat niet voldoet aan de norm waarin een gen dat wordt geërfd van een van de ouders zich afscheidt in kiembaancellen met dezelfde waarschijnlijkheid, en deze omvatten:
- Mitochondriale overerving
- Opdruk
- Uniparentale disomie
- Incomplete dominantie
- Codominantie
- Meerdere allelen
- Pleiotropie
- Dodelijke allelen
- Polygene eigenschappen
- Geslachtsgebonden overerving
Het voorkomen van deze variaties in erfelijke patronen kan worden toegeschreven aan de verschillende interacties die genen hebben met andere cellulaire componenten, naast het feit dat elk ervan onderhevig is aan regulatie en variatie in elk van de stadia van transcriptie, splitsing, translatie, proteïne. vouwen, oligomerisatie, translocatie en compartimentering binnen de cel en voor de export ervan.
Met andere woorden, er zijn talloze epigenetische invloeden die de overervingspatronen van een eigenschap kunnen wijzigen, wat resulteert in een "afwijking" van de wetten van Mendel..
Mitochondriaal DNA zendt ook informatie van de ene generatie naar de andere, net als dat wat zich in de kern van alle eukaryote cellen bevindt. Het genoom dat in dit DNA wordt gecodeerd, omvat de genen die nodig zijn voor de synthese van 13 polypeptiden die deel uitmaken van de subeenheden van de mitochondriale ademhalingsketen, essentieel voor organismen met aëroob metabolisme..
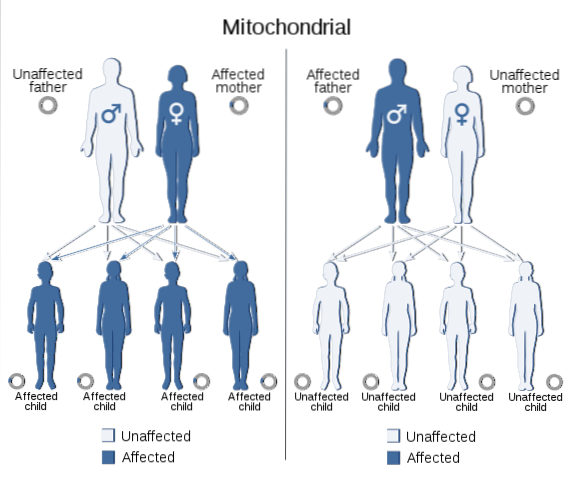
Die eigenschappen die het gevolg zijn van mutaties in het mitochondriale genoom vertonen een specifiek segregatiepatroon dat 'mitochondriale overerving' wordt genoemd, wat meestal plaatsvindt via de moederlijn, aangezien de eicel het totale complement van mitochondriaal DNA levert en er geen mitochondriën worden bijgedragen door sperma..
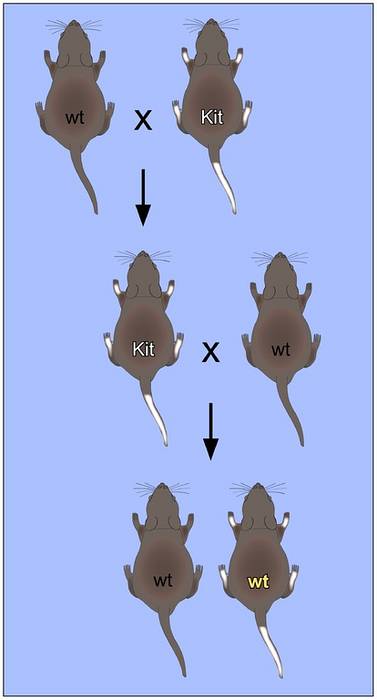
Genomische inprenting bestaat uit een reeks epigenetische "kenmerken" die bepaalde genen of complete genomische regio's karakteriseren en die het resultaat zijn van de genomische overgang van het mannetje of het vrouwtje door het proces van gametogenese..
Er zijn gen-imprinting clusters, die bestaan uit 3 tot 12 genen verdeeld over 20 tot 3700 kilo basen van DNA. Elk cluster heeft een gebied dat bekend staat als het controlegebied van de imprinting, dat specifieke epigenetische modificaties van elke ouder vertoont, waaronder:
- DNA-methylering op specifieke allelen in de cytokineresiduen van CpG-paren
- Post-translationele modificatie van chromatine-gerelateerde histonen (methylering, acetylering, fosforylering, enz., Van de aminozuurstaarten van deze eiwitten).
Beide typen "merktekens" moduleren permanent de expressie van de genen waarop ze worden aangetroffen, waardoor hun transmissiepatronen worden aangepast aan de volgende generatie..
De overervingspatronen waarin de expressie van een ziekte afhangt van de specifieke allelen die van beide ouders worden geërfd, staat bekend als het ouderlijke oorsprongseffect..
Dit fenomeen is een uitzondering op de eerste wet van Mendel, die stelt dat slechts één van de twee allelen die in elke ouder aanwezig zijn, wordt overgedragen op het nageslacht en, volgens de chromosomale wetten van overerving, slechts één van de homologe chromosomen van de ouders kan worden overgedragen. de volgende generatie.
Dit is een uitzondering op de regel, aangezien uniparentale disomie de overerving is van beide kopieën van een homoloog chromosoom van een van de ouders. Dit type overervingspatroon vertoont niet altijd fenotypische defecten, omdat het de numerieke en structurele kenmerken van diploïde chromosomen behoudt..
Dit overervingspatroon bestaat, fenotypisch gesproken, uit een mengsel van allelgecodeerde eigenschappen die worden gecombineerd. In gevallen van onvolledige dominantie vertonen die individuen die heterozygoot zijn een mengeling van de eigenschappen van de twee allelen die hen beheersen, wat impliceert dat de relatie tussen de fenotypes is gewijzigd.
Beschrijft de erfelijke patronen waarin de twee allelen die van ouders op hun kinderen worden overgedragen gelijktijdig tot uitdrukking komen in die met heterozygote fenotype, waarvoor beide als "dominant" worden beschouwd.
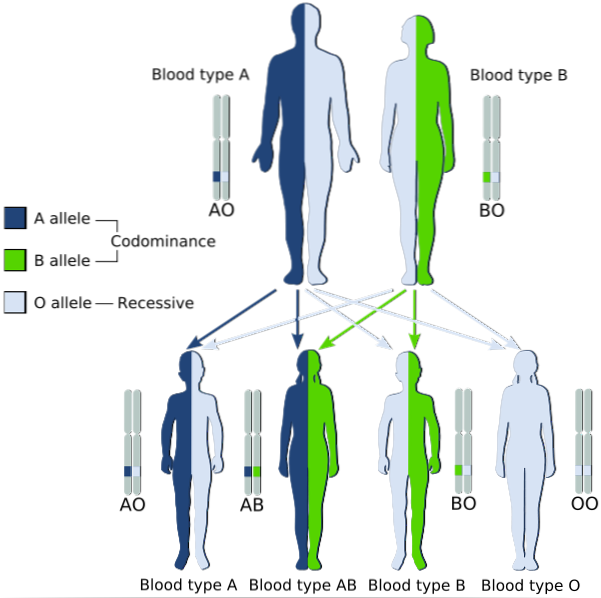
Met andere woorden, het recessieve allel wordt niet "gemaskeerd" door de expressie van het dominante allel in het allelpaar, maar beide komen tot expressie en een mengsel van de twee eigenschappen wordt waargenomen in het fenotype..
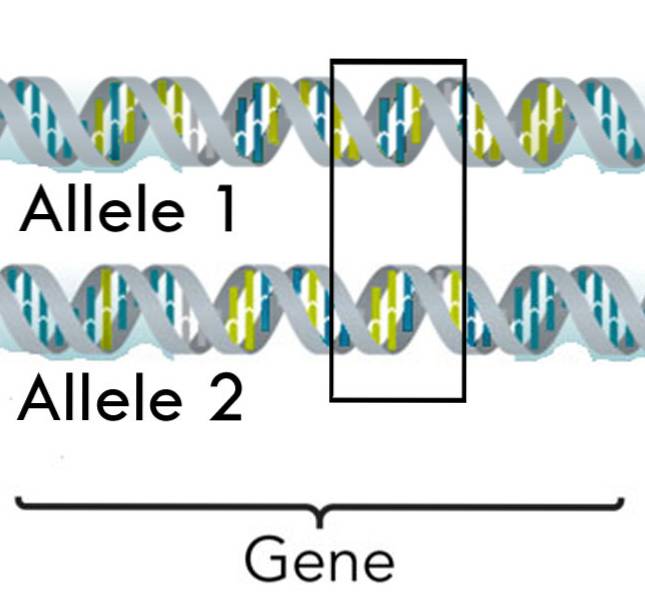
Misschien wordt een van de belangrijkste zwakheden van de Mendeliaanse overerving vertegenwoordigd door eigenschappen die worden gecodeerd door meer dan één allel, wat vrij vaak voorkomt bij mensen en veel andere levende wezens..
Dit erfelijke fenomeen vergroot de diversiteit van de eigenschappen die door een gen worden gecodeerd en bovendien kunnen deze genen naast eenvoudige of volledige dominantie ook patronen van onvolledige dominantie en codominantie ervaren..
Een andere "steen in de schoen" of "losse benen" van Mendel's erfelijke theorieën heeft te maken met die genen die het verschijnen van meer dan één zichtbaar fenotype of kenmerk controleren, zoals het geval is bij pleiotrope genen..
In zijn werken hield Mendel ook geen rekening met de overerving van bepaalde allelen die het voortbestaan van het nageslacht kunnen voorkomen wanneer het in de homozygote of heterozygote vorm is; dit zijn de dodelijke allelen.
Dodelijke allelen zijn meestal gerelateerd aan mutaties of defecten in genen die strikt noodzakelijk zijn om te overleven, die, afhankelijk van de homozygotie of heterozygositeit van de individuen, dodelijk zijn wanneer ze worden overgedragen aan de volgende generatie (dergelijke mutaties)..
Er zijn kenmerken die worden gecontroleerd door meer dan één gen (met hun allelen) en die bovendien sterk worden gecontroleerd door de omgeving. Bij mensen komt dit buitengewoon vaak voor en is het het geval voor eigenschappen zoals lengte, oog-, haar- en huidskleur, evenals het risico om aan bepaalde ziekten te lijden..
Bij mensen en veel dieren zijn er ook eigenschappen die worden aangetroffen op een van de twee geslachtschromosomen en die worden overgedragen via seksuele voortplanting. Veel van deze eigenschappen worden als "geslachtsgebonden" beschouwd wanneer ze slechts bij één van de geslachten voorkomen, hoewel beide fysiek in staat zijn om deze eigenschappen te erven..
De meeste geslachtsgebonden kenmerken worden in verband gebracht met enkele recessieve ziekten en aandoeningen.
Er is een genetische aandoening bij mensen bekend als het Marfan-syndroom, die wordt veroorzaakt door een mutatie in een enkel gen dat tegelijkertijd de groei en ontwikkeling beïnvloedt (onder andere lengte, gezichtsvermogen en hartfunctie)..
Dit is een geval dat wordt beschouwd als een uitstekend voorbeeld van het niet-Mendeliaanse overervingspatroon dat pleiotropie wordt genoemd, waarin een enkel gen verschillende kenmerken controleert..
Genetische aandoeningen die het gevolg zijn van mutaties in mitochondriaal DNA vertonen een aantal klinische fenotypische variaties, aangezien wat bekend staat als heteroplasmie optreedt, waarbij verschillende weefsels een verschillend percentage mutant mitochondriaal genoom hebben en daarom verschillende fenotypes vertonen..
Onder deze aandoeningen bevinden zich de mitochondriale "depletie" -syndromen, die een groep autosomaal recessieve aandoeningen zijn die worden gekenmerkt door een significante vermindering van het gehalte aan mitochondriaal DNA, wat eindigt met gebrekkige energieproductiesystemen in de meest getroffen organen en weefsels..
Deze syndromen kunnen het gevolg zijn van mutaties in het nucleaire genoom die de nucleaire genen beïnvloeden die betrokken zijn bij de synthese van mitochondriale nucleotiden of bij de replicatie van mitochondriaal DNA. De effecten kunnen worden aangetoond als myopathieën, encefalopathieën, hepato-cerebrale of neuro-gastro-intestinale defecten.



Niemand heeft nog op dit artikel gereageerd.