De nucleosoom het is de basiseenheid van DNA-verpakking in eukaryote organismen. Het is daarom het kleinste chromatine-compressie-element.
Het nucleosoom is gebouwd als een octameer van eiwitten die histonen worden genoemd, of een trommelvormige structuur waarop ongeveer 140 nt DNA is gewikkeld, waardoor bijna twee volledige omwentelingen worden gemaakt..
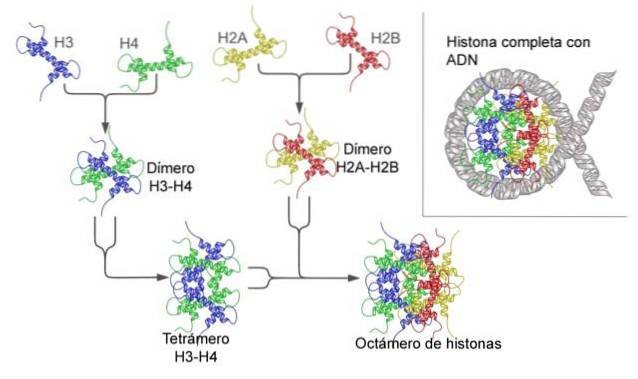
Bovendien wordt nog eens 40-80 nt DNA beschouwd als onderdeel van het nucleosoom, en het is de DNA-fractie die fysieke continuïteit mogelijk maakt tussen het ene nucleosoom en het andere in complexere chromatinestructuren (zoals de 30 nm chromatinevezel)..
De histoncode was een van de eerste moleculair begrepen epigenetische controle-elementen..
Artikel index
Nucleosomen maken het mogelijk:
In de meest fundamentele zin bestaan nucleosomen uit DNA en eiwitten. DNA kan vrijwel elk dubbelband-DNA zijn dat aanwezig is in de kern van de eukaryote cel, terwijl nucleosomale eiwitten allemaal behoren tot de reeks eiwitten die histonen worden genoemd..
Histonen zijn kleine eiwitten met een hoge belasting aan basische aminozuurresiduen; Dit maakt het mogelijk om de hoge negatieve lading van DNA tegen te gaan en een efficiënte fysische interactie tussen de twee moleculen tot stand te brengen zonder de starheid van de covalente chemische binding te bereiken..
Histonen vormen een drumachtige octameer met twee kopieën of monomeren van elk van de histonen H2A, H2B, H3 en H4. Het DNA maakt bijna twee volledige omwentelingen aan de zijkanten van de octameer en gaat dan verder met een fractie van het linker-DNA dat associeert met histon H1, om terug te keren om twee volledige omwentelingen te geven op een andere histon-octameer..
De octameer-set, het bijbehorende DNA en het bijbehorende linker-DNA is een nucleosoom.
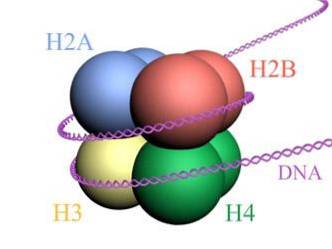
Genomisch DNA bestaat uit extreem lange moleculen (meer dan een meter in het geval van mensen, rekening houdend met al hun chromosomen), die moeten worden gecomprimeerd en georganiseerd in een extreem kleine kern.
De eerste stap in deze verdichting wordt uitgevoerd door de vorming van nucleosomen. Alleen al met deze stap wordt DNA ongeveer 75 keer gecomprimeerd.
Hierdoor ontstaat een lineaire vezel waaruit opeenvolgende niveaus van chromatine-verdichting worden opgebouwd: de 30 nm-vezel, de lussen en de lussen van lussen..
Wanneer een cel zich deelt, hetzij door mitose of door meiose, is de ultieme mate van verdichting respectievelijk het mitotische of meiose chromosoom zelf..
Het feit dat histon-octameren en DNA elektrostatisch interageren, verklaart gedeeltelijk hun effectieve associatie, zonder de vloeibaarheid te verliezen die nodig is om nucleosomen dynamische elementen te maken voor het verdichten en decompacteren van chromatine..
Maar er is een nog verrassender interactie-element: de N-terminale uiteinden van de histonen worden blootgesteld buiten het inwendige van de octameer, die compacter en inert is..
Deze uiteinden staan niet alleen fysiek in wisselwerking met het DNA, maar ondergaan ook een reeks covalente modificaties waarvan de mate van verdichting van het chromatine en de expressie van het bijbehorende DNA zal afhangen..
De verzameling covalente wijzigingen, onder meer in termen van type en aantal, wordt gezamenlijk de histoncode genoemd. Deze modificaties omvatten fosforylering, methylering, acetylering, ubiquitinering en sumoylering van arginine- en lysineresiduen aan de N-uiteinden van histonen..
Elke verandering, samen met andere binnen hetzelfde molecuul of in residuen van andere histonen, in het bijzonder histonen H3, zal de expressie of niet van het geassocieerde DNA bepalen, evenals de mate van verdichting van het chromatine..
Als algemene regel is bijvoorbeeld gezien dat gehypermethyleerde en gehypoacetyleerde histonen bepalen dat het geassocieerde DNA niet tot expressie wordt gebracht en dat chromatine in een compactere toestand aanwezig is (heterochromatisch en daarom inactief)..
Daarentegen wordt euchromatisch DNA (minder compact en genetisch actief) geassocieerd met een chromatine waarvan de histonen hyperacetyleerd en gehypomethyleerd zijn..
We hebben al gezien dat de covalente modificatiestatus van histonen de mate van expressie en lokale chromatine-verdichting kan bepalen. Op mondiaal niveau wordt chromatine-verdichting ook gereguleerd door covalente modificaties van histonen in nucleosomen.
Er is bijvoorbeeld aangetoond dat constitutief heterochromatine (dat nooit tot expressie komt en dicht opeengepakt is) de neiging heeft zich te hechten aan de nucleaire lamina, waardoor de nucleaire poriën vrij blijven..
Van zijn kant doet constitutief euchromatine (dat altijd tot expressie wordt gebracht, zoals datgene dat celonderhoudsgenen omvat en zich bevindt in gebieden van laks chromatine), dit in grote lussen die het te transcriberen DNA blootstellen aan de transcriptiemachine.
Andere regio's van genomisch DNA oscilleren tussen deze twee toestanden, afhankelijk van de ontwikkelingstijd van het organisme, groeiomstandigheden, celidentiteit, enz..
Om hun plan voor celontwikkeling, -expressie en -onderhoud te vervullen, moeten de genomen van eukaryote organismen nauwkeurig reguleren wanneer en hoe hun genetische mogelijkheden zich moeten manifesteren..
Uitgaande van de informatie die is opgeslagen in hun genen, bevinden deze zich in de kern in bepaalde regio's die hun transcriptietoestand bepalen..
We kunnen daarom zeggen dat een andere fundamentele rol van nucleosomen, door de veranderingen in chromatine die het helpt te definiëren, de organisatie of architectuur is van de kern waarin ze zijn ondergebracht..
Deze architectuur is geërfd en is fylogenetisch geconserveerd dankzij het bestaan van deze modulaire elementen van informatieve verpakkingen.



Niemand heeft nog op dit artikel gereageerd.