De G-eiwitten of guanine nucleotide bindende eiwitten, zijn eiwitten die zijn geassocieerd met het plasmamembraan en behoren tot een familie van signaalkoppelingeiwitten die belangrijke functies hebben in veel signaaltransductieprocessen in eukaryote organismen.
In de literatuur worden G-eiwitten beschreven als binaire moleculaire schakelaars, aangezien hun biologische activiteit wordt bepaald door de veranderingen in hun structuur door de nucleotidesoort waaraan ze kunnen binden: guanosinenucleotiden (difosfaat (GDP) en trifosfaat (GTP) ).
Ze worden over het algemeen geactiveerd door receptoren van een familie van eiwitten die bekend staan als G Protein Coupled Receptors (GPCR's).G-proteïne-gekoppelde receptoren "), die een extern initieel signaal ontvangen en dit omzetten in conformatieveranderingen die activering veroorzaken, wat zich vervolgens vertaalt in de activering van een ander effectoreiwit.
Sommige auteurs zijn van mening dat de coderende genen voor deze familie van eiwitten zijn geëvolueerd door duplicatie en divergentie van een gemeenschappelijk voorouderlijk gen, waarvan het product steeds verfijnder en gespecialiseerd werd..
Onder de grote verscheidenheid aan cellulaire functies die deze eiwitten hebben, zijn de translocatie van macromoleculen tijdens de eiwitsynthese, de transductie van hormonale signalen en sensorische stimuli, evenals de regulering van celproliferatie en differentiatie..
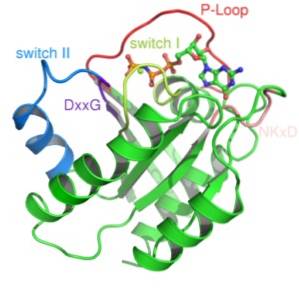
Er zijn twee klassen van dit type eiwit beschreven: kleine G-eiwitten en heterotrimere G-eiwitten. De eerste driedimensionale structuur van een G-eiwit werd meer dan tien jaar geleden verkregen uit een klein G-eiwit dat bekend staat als Ras.
Artikel index
Structureel gezien worden twee soorten G-eiwitten herkend: de kleine G-eiwitten en de veel complexere heterotrimere G-eiwitten..
Kleine G-eiwitten bestaan uit een enkel polypeptide van ongeveer 200 aminozuurresiduen en ongeveer 20-40 kDa, en in hun structuur is er een geconserveerd katalytisch domein (het G-domein) dat bestaat uit vijf α-helices, zes β-gevouwen vellen , en vijf polypeptidelussen.
Heterotrimere G-eiwitten zijn daarentegen integrale membraaneiwitten die zijn samengesteld uit drie polypeptideketens, bekend als de α-, β- en γ-subeenheden..
-De α-subeenheid weegt tussen 40 en 52 kDa, heeft een guanine-nucleotide-bindend gebied en heeft GTPase-activiteit om bindingen tussen de fosfaatgroepen van GTP te hydrolyseren..
De α-subeenheden van verschillende G-eiwitten delen enkele structurele domeinen, zoals die voor binding en hydrolyse van GTP, maar verschillen sterk in hun bindingsplaatsen voor de receptor- en effector-eiwitten..
-De β-subeenheid heeft een iets lager molecuulgewicht (tussen 35 en 36 kDa).
-De γ-subeenheid is daarentegen veel kleiner en heeft een geschat molecuulgewicht van 8 kDa..
Alle heterotrimere G-eiwitten hebben 7 transmembraandomeinen en delen sequentie-overeenkomst met de β- en γ-domeinen. Deze twee domeinen zijn zo sterk met elkaar verbonden dat ze als een enkele functionele eenheid worden beschouwd..
Zoals hierboven vermeld, zijn er twee soorten G-eiwitten: klein en heterotrimeer..
Kleine G-eiwitten spelen een rol bij celgroei, eiwitafscheiding en intracellulaire vesikelinteractie. Op hun beurt zijn heterotrimere G-eiwitten geassocieerd met de transductie van signalen van oppervlaktereceptoren en fungeren ze ook als schakelaars die tussen twee toestanden wisselen, afhankelijk van het bijbehorende nucleotide..
Deze eiwitten worden ook wel kleine GTPases, kleine GTP-bindende eiwitten of Ras-eiwit-superfamilie genoemd en vormen een onafhankelijke superfamilie binnen de grote klasse van GTP-hydrolasen met regulerende functies..
Deze eiwitten zijn zeer divers en sturen meerdere cellulaire processen aan. Ze worden gekenmerkt door een geconserveerd GTP-bindend domein, het "G" -domein. De binding van dit fosfaatnucleotide veroorzaakt belangrijke conformatieveranderingen in zijn katalytische domein in kleine G-eiwitten..
Zijn activiteit is nauw verwant aan GTPase-activerende eiwitten (GAP).GTPase Activating Protein ") en met guanine-nucleotide-uitwisselingsfactoren (GEF)Guanine Nucleotide Exchange Factor ”).
In eukaryoten zijn vijf klassen of families van kleine G-eiwitten beschreven:
-Ras
-Rho
-Rab
-Sar1 / Arf
-Ran
De Ras- en Rho-eiwitten regelen de genexpressie en de Rho-eiwitten moduleren ook de reorganisatie van het cytoskelet. De eiwitten uit de Rab- en Sar1 / Arf-groep beïnvloeden het vesiculaire transport en de Ran-eiwitten reguleren het nucleair transport en de celcyclus..
Dit type eiwit verdient ook een associatie met twee andere eiwitfactoren, zodat de signaalroute van de externe omgeving naar het binnenste van de cel bestaat uit drie elementen in de volgende volgorde:
Er is een grote diversiteit aan heterotrimere G-eiwitten en dit houdt verband met de grote diversiteit aan α-subeenheden die in de natuur voorkomen, waarin slechts 20% van de aminozuursequentie behouden is.
Gewoonlijk worden heterotrimere G-eiwitten geïdentificeerd dankzij de diversiteit van de α-subeenheid, voornamelijk gebaseerd op hun functionele en sequentieovereenkomsten..
De α-subeenheden bestaan uit vier families (de familie Gs, de familie Gi / o, de familie Gq en de familie G12). Elke familie bestaat uit een ander "isotype" dat samen meer dan 15 verschillende vormen van α-subeenheden toevoegt..
Deze familie bevat vertegenwoordigers die ook deelnemen aan de opregulatie van adenylaatcyclase-eiwitten en komt tot expressie in de meeste celtypen. Het bestaat uit twee leden: Gs en Golf.
Het abonnement 's " verwijst naar stimulatie (uit het Engels "stimulatie ') en het subscript “olf " verwijst naar "geur" (uit het Engels "Reukzin"Golfproteïnen komen vooral tot uiting in sensorische neuronen die verantwoordelijk zijn voor geur.
Dit is de grootste en meest diverse familie. Ze komen tot uiting in veel celtypen en mediëren receptorafhankelijke remming van verschillende soorten adenylcyclasen (het subscript “ik " verwijst naar remming).
Eiwitten met de α-subeenheden van de Go-groep komen vooral tot expressie in cellen van het centrale zenuwstelsel en hebben twee varianten: A en B..
Eiwitten met deze α-subeenheidfamilie zijn verantwoordelijk voor de regulatie van fosfolipase C. Deze familie bestaat uit vier leden waarvan de α-subeenheden tot expressie worden gebracht door verschillende genen. Ze zijn overvloedig aanwezig in levercellen, niercellen en longen.
Deze familie komt alomtegenwoordig tot expressie in organismen en het is niet met zekerheid bekend wat precies de cellulaire processen zijn die door eiwitten met deze subeenheden worden gereguleerd..
Β en γ subeenheden
Hoewel de diversiteit aan alfa-structuren bepalend is voor de identificatie van heterotrimere eiwitten, is er ook veel diversiteit met betrekking tot de andere twee subeenheden: bèta en gamma..
G-eiwitten nemen deel aan het "kanaliseren" van signalen van receptoren op het plasmamembraan naar kanalen of effectorenzymen..
Het meest voorkomende voorbeeld van de functie van dit type eiwit is de regulering van het enzym adenylaatcyclase, een enzym dat verantwoordelijk is voor de synthese van adenosine 3 ', 5'-monofosfaat of simpelweg cyclisch AMP, een molecuul dat belangrijke functies heeft als een tweede boodschapper in veel bekende cellulaire processen:
-Selectieve fosforylering van eiwitten met specifieke functies
-Genetische transcriptie
-Reorganisatie van het cytoskelet
-Afscheiding
-Membraan depolarisatie
Ze nemen ook indirect deel aan de regulering van de inositol-signaalcascade (fosfatidylinositol en zijn gefosforyleerde derivaten), die verantwoordelijk zijn voor de controle van calciumafhankelijke processen zoals chemotaxis en de uitscheiding van oplosbare factoren..
Veel ionenkanalen en transporteiwitten worden direct aangestuurd door eiwitten van de G-eiwitfamilie. Op dezelfde manier zijn deze eiwitten betrokken bij vele sensorische processen zoals zicht, reuk en andere..
De wijze van interactie van een G-eiwit met de effector-eiwitten is specifiek voor elke klasse of familie van eiwitten.
Voor G-eiwitten gekoppeld aan membraanreceptoren (heterotrimere G-eiwitten) veroorzaakt de binding van een guaninenucleotide zoals GDP of guanosinedifosfaat aan de α-subeenheid de associatie van de drie subeenheden, waardoor een complex wordt gevormd dat bekend staat als Gαβγ of G-GDP, dat bevestigd aan het membraan.
Als het GDP-molecuul vervolgens wordt vervangen door een GTP-molecuul, dissocieert de α-subeenheid die aan GTP is gehecht van de β- en γ-subeenheden, waardoor een afzonderlijk complex wordt gevormd dat bekend staat als Gα-GTP, dat in staat is om de activiteit van zijn enzymen of doeldrager-eiwitten te veranderen.
De hydrolytische activiteit van deze subeenheid stelt het in staat de activering te beëindigen, de GTP in te ruilen voor een nieuw GDP en over te gaan naar de inactieve conformatie..
Bij afwezigheid van de aangeslagen receptoren die associëren met G-eiwitten, is dit proces van uitwisseling van GDP naar GTP erg traag, wat betekent dat heterotrimere G-eiwitten alleen GDP voor GTP uitwisselen met een fysiologisch significante snelheid wanneer ze gebonden zijn aan hun aangeslagen receptoren..



Niemand heeft nog op dit artikel gereageerd.