
De genetische recombinatie het is het proces waarbij nucleïnezuurmoleculen fragmenten uitwisselen, waardoor een nieuw molecuul ontstaat. Het komt veel voor in DNA, maar RNA is ook een substraat voor recombinatie. Recombinatie is, na mutatie, de belangrijkste bron voor het genereren van genetische variabiliteit.
DNA neemt deel aan verschillende biochemische processen. Tijdens replicatie dient het als een sjabloon voor het genereren van twee nieuwe DNA-moleculen. Bij transcriptie maakt het de generatie mogelijk van RNA-moleculen uit specifieke regio's die worden gecontroleerd door een promotor.
Maar daarnaast is DNA ook in staat fragmenten uit te wisselen. Door dit proces genereert het nieuwe combinaties die niet het product zijn van de twee voorgaande processen, noch van bemesting..
Alle recombinatieprocessen omvatten het breken en samenvoegen van de DNA-moleculen die aan het proces deelnemen. Dit mechanisme varieert afhankelijk van het recombinatiesubstraat, de enzymen die deelnemen aan het proces en het mechanisme van de uitvoering ervan..
Recombinatie hangt in het algemeen af van het bestaan van complementaire, vergelijkbare (zo niet identieke) of homologe gebieden tussen de recombinerende moleculen. In het geval dat moleculen recombineren in processen die niet door homologie worden geleid, wordt gezegd dat de recombinatie niet-homoloog is.
Als de homologie betrekking heeft op een zeer kort gebied dat in beide moleculen aanwezig is, wordt gezegd dat de recombinatie plaatsspecifiek is..
Artikel index
Wat we homologie in recombinatie noemen, verwijst niet noodzakelijk naar de evolutionaire oorsprong van de deelnemende moleculen. We hebben het meer over de mate van overeenkomst in de nucleotidesequentie.
Niet-herstellende recombinatie komt bijvoorbeeld voor in eukaryoten tijdens het meioseproces. Er kan ongetwijfeld geen grotere homologie zijn dan tussen chromosomenparen in dezelfde cel.
Daarom worden ze homologe chromosomen genoemd. Er zijn echter gevallen waarin het DNA van een cel materiaal uitwisselt met een vreemd DNA. Deze DNA's moeten erg lijken op recombineren, maar ze hoeven niet noodzakelijk dezelfde voorouder (homologie) te delen om dit te bereiken..
De plaats van aanhechting en uitwisseling tussen twee DNA-moleculen wordt het chiasma genoemd en het proces zelf wordt verknoping genoemd. Tijdens de cross-over wordt een uitwisseling van banden geverifieerd tussen de deelnemende DNA's..
Dit genereert een co-integratie, twee DNA-moleculen die fysiek tot één zijn samengevoegd. Bij het "oplossen" van het cointegrate, worden twee moleculen gegenereerd, over het algemeen veranderd (recombinant).
"Oplossen", in de context van recombinatie, is het scheiden van de samenstellende DNA-moleculen van een co-integratie.
Bij plaatsspecifieke recombinatie hebben twee DNA-moleculen, in het algemeen niet homoloog, een korte sequentie die beide gemeen hebben. Deze sequentie is het doelwit van een specifiek splitsingsenzym.
Het enzym, dat deze sequentie kan herkennen en niet een andere, snijdt het op een bepaalde plaats in beide moleculen. Met behulp van een aantal andere factoren, wisselt het de DNA-banden van de twee deelnemende moleculen uit en vormt het een co-integratie.
Dit is de basis van de vorming van het cointegrate tussen het genoom van de bacterie Escherichia coli en die van de bacteriofaag lambda. Een bacteriofaag is een virus dat bacteriën infecteert.
De vorming van dit cointegraat wordt uitgevoerd door een enzym dat wordt gecodeerd in het virusgenoom: lambda integrase. Het herkent een gemeenschappelijke reeks genaamd attP in het circulaire genoom van het virus, en attB in dat van de bacteriën.
Door beide sequenties in beide moleculen te knippen, genereert het vrije segmenten, verwisselt het de banden en voegt het zich bij de twee genomen. Er wordt dan een grotere of samengeïntegreerde cirkel gevormd.
Bij co-integratie wordt het virusgenoom passief gedragen door het bacteriële genoom, waarmee het repliceert. In deze toestand wordt gezegd dat het virus zich in de provirustoestand bevindt en dat de bacterie er lysogeen voor is..
Het omgekeerde proces, dat wil zeggen de oplossing van de co-integratie, kan vele generaties duren - of zelfs niet plaatsvinden. Dit wordt echter enzymatisch gemedieerd door een ander eiwit dat wordt gecodeerd door het virusgenoom, excisionase genaamd. Wanneer dit gebeurt, scheidt het virus zich van de co-integratie, reactiveert het en veroorzaakt cellysis..
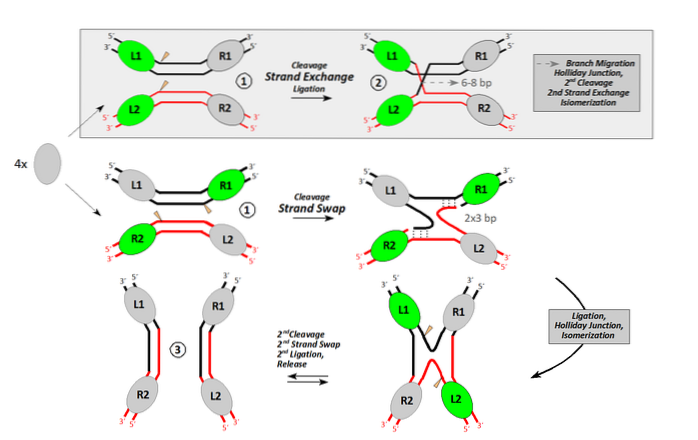
Homologe recombinatie vindt plaats tussen DNA-moleculen die ten minste ongeveer 40 nucleotiden met volledige of bijna volledige gelijkenis delen. Om het recombinatieproces uit te voeren, moet ten minste één endonuclease deelnemen.
Endonucleasen zijn enzymen die interne sneden in het DNA maken. Sommigen doen het om door te gaan met het afbreken van DNA. Anderen, zoals in het geval van recombinatie, doen het om een deuk in het DNA te genereren.
Deze unieke nick maakt het mogelijk om een enkelband-DNA met een vrij uiteinde te verwerken. Dit vrije uiteinde, georiënteerd door een recombinase, laat een enkele band toe om een dubbel DNA binnen te dringen, waardoor de residente band identiek eraan wordt verplaatst..
Dit is het kruispunt tussen een donor ("indringer") DNA-molecuul en een andere receptor..
Het enzym (recombinase) dat het proces van bandinvasie en -uitwisseling uitvoert Escherichia coli het heet RecA. Er zijn andere homologe eiwitten in prokaryoten, zoals RadA in archaea. In eukaryoten wordt het equivalente enzym RAD51 genoemd.
Zodra de invasieve band de bewoner verplaatst, werkt ze samen met de band die eenvoudig in het donormolecuul is achtergelaten. Beide plaatsen worden afgesloten door de werking van een ligase.
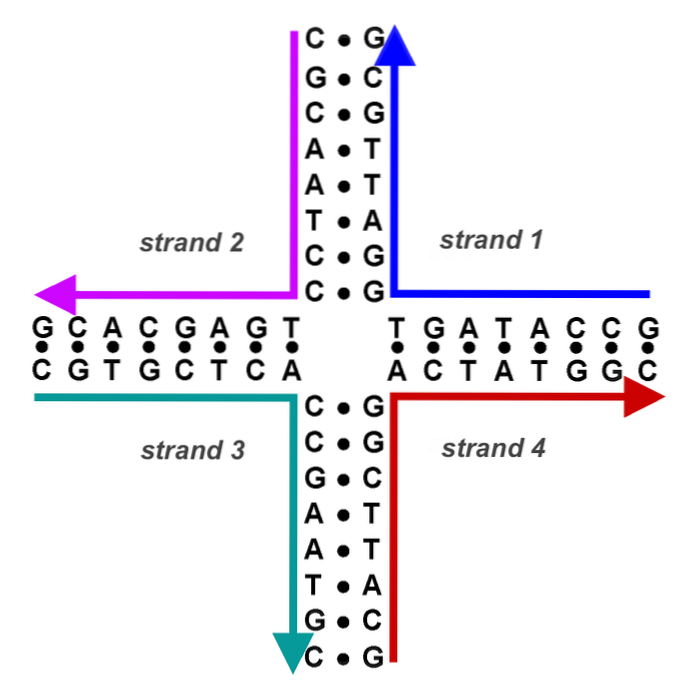
Nu hebben we een DNA van hybride banden (een donorband en een ontvangende band, van verschillende oorsprong) geflankeerd door donor-DNA en ontvanger-DNA. De kruispunten (chiasmata) bewegen in beide richtingen met minstens 200 bp.
Elk kruispunt vormt de zogenaamde Holliday-structuur (kruisvormig DNA-product van een recombinatiegebeurtenis).
Dit kruisvormige DNA moet worden gescheiden door andere endonucleasen. Chimerisch of hybride DNA van deze structuur kan op twee manieren worden opgelost. Als de tweede endonucleotide-cut plaatsvindt in dezelfde band als de eerste, vindt geen recombinatie plaats. Als de tweede snede plaatsvindt in de andere band, zijn de resulterende producten recombinant.
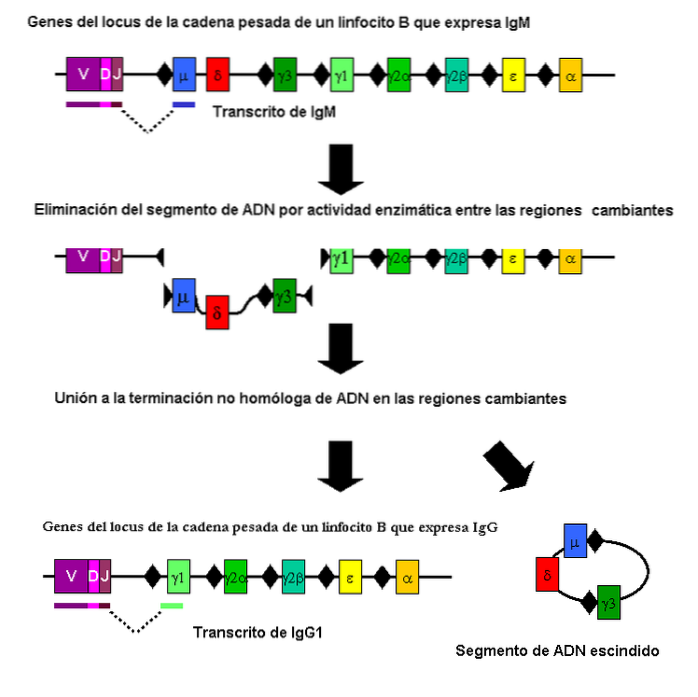
Dit is een soort somatische recombinatie (niet meiotisch) die bijdraagt aan het genereren van de enorme variabiliteit van antilichamen van het immuunsysteem.
Deze recombinatie vindt plaats in bepaalde fragmenten van de genen die coderen voor de polypeptideketens die ze definiëren. Het wordt uitgevoerd door B-cellen en omvat verschillende genetische regio's.
Interessant is dat er parasieten zijn zoals Trypanosoma brucei die een soortgelijk recombinatiemechanisme gebruiken om variabiliteit in een oppervlakteantigeen te creëren. Op deze manier kunnen ze de reactie van de gastheer omzeilen als de gastheer er niet in slaagt het antilichaam te genereren dat het "nieuwe" antigeen kan herkennen..
Ten slotte zijn er recombinatieprocessen die niet afhankelijk zijn van de gelijkenis in de volgorde van de deelnemende moleculen. In eukaryoten is bijvoorbeeld de recombinatie van niet-homologe uiteinden erg belangrijk.
Dit gebeurt bij DNA-fragmenten die dubbele bandbreuken in het DNA vertonen. Deze worden "gerepareerd" doordat de cel ze met andere fragmenten verbindt, ook met dubbele bandbreuken.
Deze moleculen hoeven echter niet per se vergelijkbaar te zijn om deel te nemen aan dit recombinatieproces. Dat wil zeggen, door de schade te herstellen, kan de cel niet-verwante DNA's samenvoegen, waardoor een echt nieuw (recombinant) molecuul ontstaat.
Recombinatie zorgt voor de getrouwheid van DNA-informatie tijdens en na het replicatieproces. Recombinatie detecteert DNA-schade tijdens het nieuwe bandvormingsproces in dit extreem lange macromolecuul.
Aangezien elke band zijn eigen informatie heeft, en die van zijn complementaire, garandeert recombinatie dat er geen verloren gaat. Elk treedt op als getuige voor de ander. Evenzo is in diploïde organismen een homoloog chromosoom getuige van zijn broer of zus, en vice versa.
Aan de andere kant, zodra DNA is gerepliceerd, zijn de schadeherstelmechanismen van dit molecuul gevarieerd. Sommige zijn direct (er wordt direct op gehandeld) en andere zijn indirect.
Indirecte herstelmechanismen zijn afhankelijk van het plaatsvinden van recombinatie. Dat wil zeggen, om de schade in een DNA-molecuul te herstellen, wordt een ander homoloog molecuul gebruikt. Dit zou fungeren in herstellende recombinatie als een sjabloon waaraan het schade heeft geleden..
Recombinatie is in staat om tijdens de meiose een enorme chromosomale variabiliteit te creëren. Somatische recombinatie genereert ook variabiliteit, zoals in het geval van antilichamen bij gewervelde dieren.
In veel organismen is meiose gametisch. Bij seksueel gereproduceerde organismen is recombinatie een van de krachtigste manieren om variabiliteit te genereren.
Dat wil zeggen, aan spontane mutatie en chromosoomsegregatie moet recombinatie worden toegevoegd als een ander element dat gametische variabiliteit genereert..
De integratie van bacteriofaaggenomen door plaatsspecifieke recombinatie heeft daarentegen bijgedragen tot de hermodellering van het genoom van hun gastheerbacteriën..
Dit heeft bijgedragen aan het genereren van genomische variabiliteit en evolutie van deze belangrijke groep levende wezens..
We hebben al gezien dat DNA kan worden gerepareerd, maar niet wat het beschadigt. In werkelijkheid kan bijna alles het DNA beschadigen, te beginnen met een foutieve replicatie die niet wordt gecorrigeerd..
Maar daarnaast kan DNA worden beschadigd door UV-licht, ioniserende straling, vrije zuurstofradicalen door cellulaire ademhaling en wat we eten, roken, ademen, inslikken of aanraken..
Gelukkig hoef je het leven niet op te geven om DNA te beschermen. Bepaalde dingen moeten worden opgegeven, maar het grote werk wordt door de cel zelf gedaan. Deze mechanismen voor het opsporen en herstellen van schade aan DNA hebben duidelijk een genetische basis, en hun tekortkoming, enorme gevolgen.
Ziekten die verband houden met defecten in homologe recombinatie omvatten bijvoorbeeld Bloom- en Werner-syndromen, familiaire borst- en eierstokkanker, enz..



Niemand heeft nog op dit artikel gereageerd.