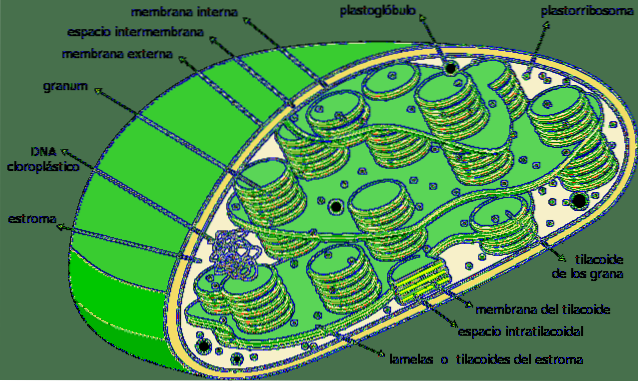
De thylakoïden zijn compartimenten in de vorm van platte zakjes in chloroplasten in plantencellen van planten, in cyanobacteriën en algen. Ze zijn meestal georganiseerd in een structuur die grana-meervoud wordt genoemd granum- en het ziet eruit als een stapel munten.
Thylakoïden worden beschouwd als het derde membraansysteem van chloroplasten, afgezien van het binnen- en buitenmembraan van het organel. Het membraan van deze structuur scheidt het inwendige van de thylakoïde van het stroma van de chloroplast en heeft een reeks pigmenten en eiwitten die betrokken zijn bij metabole routes..
In thylakoïden zijn er biochemische reacties die essentieel zijn voor fotosynthese, een proces waarbij planten zonlicht opnemen en omzetten in koolhydraten. Concreet hebben ze de nodige machines verankerd aan hun membraan om de zonne-afhankelijke fase uit te voeren, waarin licht wordt ingevangen en omgezet in energie (ATP) en NADPH..
Artikel index
Thylakoïden zijn een intern driedimensionaal vliezig systeem van chloroplasten. Volledig volgroeide chloroplasten hebben 40 tot 60 gestapelde korrels, met een diameter tussen 0,3 en 0,6 µm..
Het aantal thylakoïden waaruit spruiten bestaan, varieert sterk: van minder dan 10 zakjes in planten die aan voldoende zonlicht worden blootgesteld, tot meer dan 100 thylakoïden in planten die in extreem schaduwrijke omgevingen leven..
De gestapelde thylakoïden zijn met elkaar verbonden en vormen een continu compartiment binnen de chloroplast. Het interieur van de thylakoïde is een vrij ruim compartiment met een waterig karakter..
Het thylakoïdmembraan is essentieel voor fotosynthese, aangezien daar de eerste fase van het proces plaatsvindt.
Thylakoïden zijn de dominante structuren binnen een volledig volwassen chloroplast. Als een chloroplast wordt gevisualiseerd in de traditionele lichtmicroscoop, kunnen sommige soorten korrels worden waargenomen.
Dit zijn de thylakoïde stapels; Om deze reden noemden de eerste waarnemers van deze structuren ze "grana".
Met behulp van de elektronenmicroscoop kon het beeld worden vergroot en er werd geconcludeerd dat de aard van deze korrels eigenlijk gestapelde thylakoïden waren.
De vorming en structuur van het thylakoïdmembraan hangt af van de vorming van de chloroplast uit een nog ongedifferentieerd plastide, bekend als het protoplastide. De aanwezigheid van licht stimuleert de omzetting naar chloroplasten en later de vorming van gestapelde thylakoïden.
Bij chloroplasten en cyanobacteriën staat het thylakoïdmembraan niet in contact met het binnenste gedeelte van het plasmamembraan. De vorming van thylakoïdmembraan begint echter met invaginatie van het binnenmembraan..
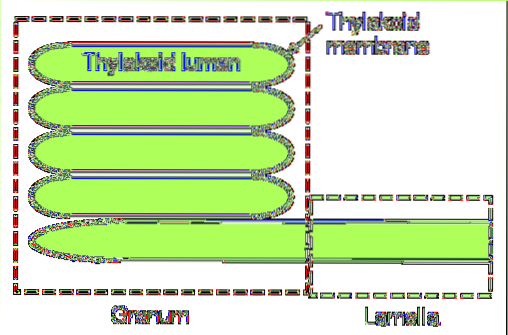
In cyanobacteriën en bepaalde soorten algen bestaan thylakoïden uit een enkele laag lamellen. Daarentegen is er een complexer systeem dat wordt aangetroffen in volwassen chloroplasten..
In deze laatste groep zijn twee essentiële onderdelen te onderscheiden: de grana en de lamel van de stroma. De eerste bestaat uit kleine gestapelde schijven en de tweede is verantwoordelijk voor het met elkaar verbinden van deze stapels, waardoor een continue structuur ontstaat: het lumen van de thylakoïde..
De lipiden waaruit het membraan bestaat, zijn zeer gespecialiseerd en bestaan voor bijna 80% uit galactosyldiacylglycerol: monogalactosyldiacylglycerol en digalactosyldiacylglycerol. Deze galactolipiden hebben sterk onverzadigde ketens, typisch voor thylakoïden..
Evenzo bevat het thylakoïdmembraan minder lipiden zoals fosfatidylglycerol. De genoemde lipiden zijn niet homogeen verdeeld in beide lagen van het membraan; er is een zekere mate van asymmetrie die lijkt bij te dragen aan het functioneren van de structuur.
Fotosystemen I en II zijn de dominante eiwitcomponenten in dit membraan. Ze zijn geassocieerd met het cytochroom b-complex6F- en ATP-synthetase.
Het is gebleken dat de meeste elementen van fotosysteem II zich in gestapelde scharlakenrode membranen bevinden, terwijl fotosysteem I zich meestal in niet-gestapelde thylakoïdmembranen bevindt. Dat wil zeggen, er is een fysieke scheiding tussen de twee fotosystemen..
Deze complexen omvatten integrale membraaneiwitten, perifere eiwitten, cofactoren en een verscheidenheid aan pigmenten..
Het inwendige van de thylakoïde bestaat uit een dikke, waterige substantie, waarvan de samenstelling verschilt van die van het stroma. Neemt deel aan fotofosforylering en slaat de protonen op die de proton-aandrijfkracht voor de synthese van ATP zullen genereren. In dit proces kan de pH van het lumen 4 bereiken.
In het lumen proteoom van het modelorganisme Arabidopsis thaliana er zijn meer dan 80 eiwitten geïdentificeerd, maar hun functies zijn niet volledig opgehelderd.
Lumenproteïnen zijn betrokken bij de regulatie van thylakoïde biogenese en bij de activiteit en omzet van proteïnen die fotosynthetische complexen vormen, vooral fotosysteem II en NAD (P) H dehydrogenase..
Het fotosyntheseproces, essentieel voor planten, begint in de thylakoïden. Het membraan dat ze begrenst met het chloroplast-stroma heeft alle enzymatische machinerie die nodig is om fotosynthetische reacties te laten plaatsvinden..
Fotosynthese kan worden onderverdeeld in twee hoofdfasen: lichtreacties en donkere reacties.
Zoals de naam al aangeeft, kunnen reacties die tot de eerste groep behoren alleen plaatsvinden in aanwezigheid van licht, terwijl die van de tweede groep met of zonder licht kunnen ontstaan. Merk op dat het niet nodig is dat de omgeving "donker" is, het is alleen onafhankelijk van het licht.
De eerste groep reacties, de ‘lichte’ reacties, komt voor in de thylakoïde en kan als volgt worden samengevat: licht + chlorofyl + 12 HtweeO + 12 NADP+ + 18 ADP + 18 Blzik à 6 Otwee + 12 NADPH + 18 ATP.
De tweede groep reacties vindt plaats in het chloroplast-stroma en neemt ATP en NADPH op die in de eerste fase zijn gesynthetiseerd om koolstof van kooldioxide naar glucose te verminderen (C6H.12OF6De tweede fase kan worden samengevat als: 12 NADPH + 18 ATP + 6 COtwee à C6H.12OF6 + 12 NADP+ + 18 ADP + 18 Blzik + 6 uurtweeOF.
Lichtreacties omvatten een reeks structuren die bekend staan als fotosystemen, die worden aangetroffen in het thylakoïdmembraan en zo'n 300 pigmentmoleculen bevatten, waaronder chlorofyl..
Er zijn twee soorten fotosystemen: het eerste heeft een maximale lichtabsorptiepiek van 700 nanometer en staat bekend als P700, terwijl de tweede P wordt genoemd680. Beide zijn ingebed in het thylakoïdmembraan.
Het proces begint wanneer een van de pigmenten een foton absorbeert en dit "stuitert" naar andere pigmenten. Wanneer een chlorofylmolecuul licht absorbeert, springt een elektron eruit en een ander molecuul absorbeert het. Het molecuul dat het elektron heeft verloren, is nu geoxideerd en heeft een negatieve lading.
De P680 vangt lichtenergie op uit chlorofyl a. In dit fotosysteem wordt een elektron in een hoger energiesysteem geworpen dan een primaire elektronenacceptor..
Dit elektron valt in fotosysteem I en gaat door de elektronentransportketen. Dit systeem van oxidatie- en reductiereacties is verantwoordelijk voor het overbrengen van protonen en elektronen van het ene molecuul naar het andere..
Met andere woorden, er is een stroom van elektronen van water naar fotosysteem II, fotosysteem I en NADPH..
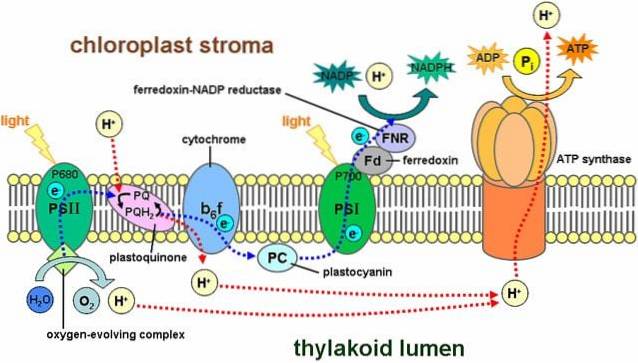
Een deel van de protonen die door dit reactiesysteem worden gegenereerd, bevindt zich in de thylakoïde (ook wel thylakoïde-licht genoemd), waardoor een chemische gradiënt ontstaat die een proton-aandrijfkracht genereert..
Protonen verplaatsen zich van de thylakoïde ruimte naar het stroma en volgen de elektrochemische gradiënt gunstig; dat wil zeggen, ze komen uit de thylakoïde.
De doorgang van protonen vindt echter niet plaats via een plaats in het membraan, ze moeten dit doen via een complex enzymatisch systeem dat ATP-synthetase wordt genoemd..
Deze beweging van protonen naar het stroma veroorzaakt de vorming van ATP vanuit ADP, een proces dat analoog is aan het proces dat plaatsvindt in de mitochondriën. De synthese van ATP met behulp van licht wordt fotofosforylering genoemd.
Deze genoemde stadia vinden gelijktijdig plaats: het chlorofyl van fotosysteem II verliest een elektron en moet dit vervangen door een elektron door de afbraak van een watermolecuul; fotosysteem I vangt licht op, oxideert en laat een elektron vrij dat wordt gevangen door NADP+.
Het verloren elektron uit fotosysteem I wordt vervangen door degene die het resultaat is van fotosysteem II. Deze verbindingen zullen worden gebruikt in de daaropvolgende koolstoffixatiereacties, in de Calvin-cyclus..
De evolutie van fotosynthese als een zuurstofafgifteproces maakte leven mogelijk zoals we dat kennen.
Er wordt voorgesteld dat fotosynthese zich enkele miljarden jaren geleden ontwikkelde in de voorouder die aanleiding gaf tot de huidige cyanobacteriën, vanuit een anoxisch fotosynthetisch complex..
Er wordt voorgesteld dat de evolutie van fotosynthese gepaard ging met twee onmisbare gebeurtenissen: de creatie van fotosysteem P680 en het ontstaan van een intern membraansysteem, zonder verbinding met het celmembraan.
Er is een eiwit genaamd Vipp1 dat essentieel is voor de vorming van thylakoïden. Dit eiwit is inderdaad aanwezig in planten, algen en cyanobacteriën, maar afwezig in bacteriën die anoxische fotosynthese uitvoeren..
Aangenomen wordt dat dit gen zijn oorsprong kan hebben in genduplicatie in de mogelijke voorouder van cyanobacteriën. Er is slechts één geval van cyanobacteriën dat in staat is tot fotosynthese met zuurstof en geen thylakoïden heeft: de soort Gloeobacter violaceus.



Niemand heeft nog op dit artikel gereageerd.