
De Golgi-apparaat, Ook bekend als Golgi-complex, het is een membraanachtig celorganel dat wordt gevormd door een reeks platte blaasjes die op elkaar zijn gestapeld; Deze zakjes bevatten vloeistof van binnen. Komt voor in een grote verscheidenheid aan eukaryoten, waaronder dieren, planten en schimmels.
Dit organel is verantwoordelijk voor de verwerking, verpakking, classificatie, distributie en modificatie van eiwitten. Daarnaast speelt het ook een rol bij de synthese van lipiden en koolhydraten. Aan de andere kant vindt in planten de synthese van de componenten van de celwand plaats in het Golgi-apparaat..
Het Golgi-apparaat werd ontdekt in 1888, terwijl zenuwcellen werden bestudeerd; zijn ontdekker, Camillo Golgi, won de Nobelprijs. De structuur is detecteerbaar door kleuring met zilverchromaat.
Aanvankelijk was het bestaan van het orgel twijfelachtig voor de wetenschappers van die tijd en zij schreven de Golgi-waarnemingen toe aan eenvoudige artefacten, producten van de gebruikte technieken.
Artikel index
Het Golgi-apparaat is een eukaryotisch organel van vliezige aard. Het lijkt op stapels zakken, hoewel de organisatie kan variëren afhankelijk van het celtype en het organisme. Het is verantwoordelijk voor de modificatie van eiwitten na translatie.
Er kan bijvoorbeeld wat koolhydraten worden toegevoegd om een glycoproteïne te vormen. Dit product wordt verpakt en gedistribueerd naar het celcompartiment waar het nodig is, zoals het membraan, lysosomen of vacuolen; het kan ook buiten de cel worden verzonden. Het neemt ook deel aan de synthese van biomoleculen.
Het cytoskelet (in het bijzonder actine) bepaalt de locatie, en over het algemeen bevindt het complex zich in een gebied van het inwendige van de cel dicht bij de kern en het centrosoom.
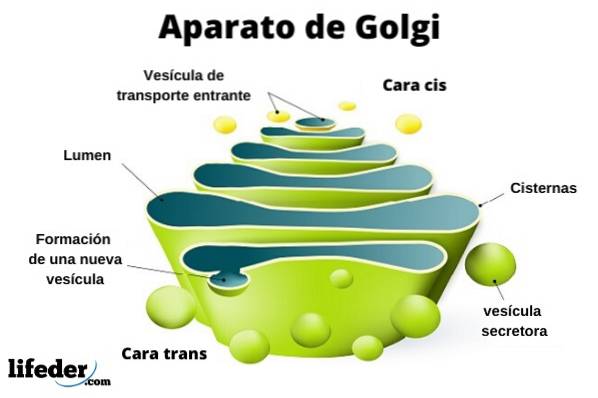
Het Golgi-complex bestaat uit een reeks platte, vensterde, schijfvormige zakjes, Golgian-stortbakken genaamd, van variabele dikte.
Deze zakken zijn gestapeld, in groepen van vier of zes reservoirs. In een zoogdiercel vind je tussen de 40 en 100 met elkaar verbonden batterijen.
Het Golgi-complex vertoont een interessant kenmerk: er is polariteit in termen van structuur en ook van functie.
U kunt onderscheid maken tussen het cis-gezicht en het trans-gezicht. De eerste houdt verband met het binnendringen van eiwitten en bevindt zich nabij het endoplasmatisch reticulum. De tweede is het uitgangsvlak of de productafscheiding; Ze worden gevormd door een of twee tanks die buisvormig zijn.
Samen met deze structuur zijn blaasjes die het transportsysteem vormen. De stapels zakken zijn aan elkaar vastgemaakt in een structuur die doet denken aan een boog of datum.
Bij zoogdieren wordt het Golgi-complex tijdens celdelingsprocessen gefragmenteerd in verschillende blaasjes. De blaasjes gaan naar de dochtercellen en nemen opnieuw de traditionele vorm van het complex aan.
De organisatie van het complex is niet bij alle groepen organismen gebruikelijk. In sommige celtypen is het complex niet gestructureerd als sets van in groepen gestapelde reservoirs; integendeel, ze bevinden zich afzonderlijk. Een voorbeeld van deze organisatie is die van de schimmel Saccharomyces cerevisiae.
In sommige eencellige organismen, zoals toxoplasma of trypanosoma, de aanwezigheid van slechts één vliezige pool is gemeld.
Al deze uitzonderingen geven aan dat het stapelen van de constructies niet essentieel is om hun functie te vervullen, hoewel de nabijheid tussen de zakken het transportproces veel efficiënter maakt..
Evenzo missen sommige basale eukaryoten deze cisternae; bijvoorbeeld paddenstoelen. Dit bewijs ondersteunt de theorie dat het apparaat in een latere lijn verscheen dan de eerste eukaryoten..
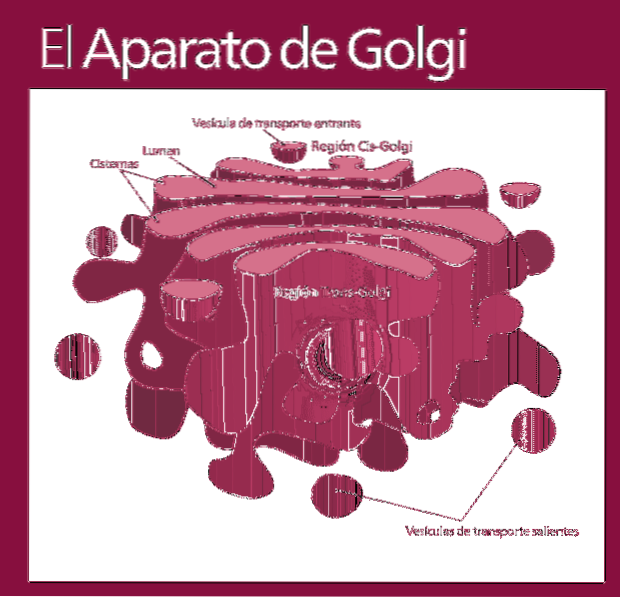
Functioneel is het Golgi-complex verdeeld in de volgende compartimenten: het cis-netwerk, de gestapelde zakjes - die op hun beurt zijn verdeeld in het middelste en trans-subcompartiment - en de trans.
De moleculen die zullen worden gemodificeerd, komen het Golgi-complex binnen in dezelfde volgorde (cis-netwerk, gevolgd door subcompartimenten die uiteindelijk worden uitgescheiden in het trans-netwerk).
De meeste reacties vinden plaats in de meest actieve zones: de trans- en middelste subcompartimenten.
Het Golgi-complex heeft als belangrijkste functie de posttranslationele modificatie van eiwitten dankzij de enzymen die ze binnenin hebben.
Deze modificaties omvatten processen van glycosylering (toevoeging van koolhydraten), fosforylering (toevoeging van een fosfaatgroep), sulfatering (toevoeging van een fosfaatgroep) en proteolyse (afbraak van eiwitten).
Bovendien is het Golgi-complex betrokken bij de synthese van specifieke biomoleculen. Elk van zijn functies wordt hieronder in detail beschreven:
In het Golgi-apparaat vindt de modificatie van een eiwit tot een glycoproteïne plaats. De typische zure pH van het inwendige van het organel is van cruciaal belang om dit proces normaal te laten verlopen..
Er is een constante uitwisseling van materialen tussen het Golgi-apparaat met het endoplasmatisch reticulum en de lysosomen. In het endoplasmatisch reticulum ondergaan eiwitten ook modificaties; deze omvatten de toevoeging van een oligosaccharide.
Wanneer deze moleculen (N-oligosacchariden) het Golgi-complex binnendringen, krijgen ze een reeks aanvullende modificaties. Als het lot van dit molecuul buiten de cel moet worden gedragen of in het plasmamembraan moet worden ontvangen, vinden er speciale wijzigingen plaats.
Deze wijzigingen omvatten de volgende stappen: de verwijdering van drie mannoseresiduen, de toevoeging van N-acetylglucosamine, de verwijdering van twee mannosen en de toevoeging van fucose, twee extra N-acetylglucosamine-, drie galactose- en drie siaalzuurresiduen..
Daarentegen worden de eiwitten die bestemd zijn voor lysosomen op de volgende manier gemodificeerd: er is geen verwijdering van mannosen als eerste stap; in plaats daarvan vindt fosforylering van deze residuen plaats. Deze stap vindt plaats in het cis-gebied van het complex.
Vervolgens worden de N-acetylglucosaminegroepen verwijderd, waarbij de mannosen achterblijven met het fosfaat dat aan het oligosaccharide wordt toegevoegd. Deze fosfaten geven aan dat het eiwit specifiek gericht moet zijn op lysosomen..
De receptoren die verantwoordelijk zijn voor het herkennen van de fosfaten die hun intracellulaire lot aangeven, bevinden zich in het trans-netwerk.
In het Golgi-complex vindt de synthese van glycolipiden en sfingomyeline plaats, waarbij ceramide (eerder gesynthetiseerd in het endoplasmatisch reticulum) als het oorspronkelijke molecuul wordt gebruikt. Dit proces is in strijd met dat van de rest van de fosfolipiden die het plasmamembraan vormen en die zijn afgeleid van glycerol..
Sfingomyeline is een klasse van sfingolipiden. Het is een overvloedig bestanddeel van zoogdiermembranen, met name zenuwcellen, waar ze deel uitmaken van de myeline-omhulling.
Na hun synthese worden ze getransporteerd naar hun uiteindelijke locatie: het plasmamembraan. Hun poolkoppen bevinden zich aan de buitenkant van het celoppervlak; deze elementen spelen een specifieke rol in celherkenningsprocessen.
In plantencellen draagt het Golgi-apparaat bij aan de synthese van de polysacchariden die de celwand vormen, met name hemicellulose en pectines. Door vesiculair transport worden deze polymeren buiten de cel gedragen.
In groenten is deze stap cruciaal en ongeveer 80% van de activiteit van het reticulum wordt toegewezen aan de synthese van polysacchariden. In feite zijn honderden van deze organellen gerapporteerd in plantencellen..
De verschillende biomoleculen - eiwitten, koolhydraten en lipiden - worden door het Golgi-complex naar hun cellulaire bestemmingen overgebracht. Eiwitten hebben een soort "code" die verantwoordelijk is voor het informeren van de bestemming waartoe het behoort.
Ze worden getransporteerd in blaasjes die het trans-netwerk verlaten en naar het specifieke celcompartiment reizen.
Eiwitten kunnen via een specifieke constitutieve route naar het membraan worden gedragen. Daarom is er een continue opname van eiwitten en lipiden in het plasmamembraan. Eiwitten waarvan de uiteindelijke bestemming het Golgi-complex is, worden hierdoor vastgehouden.
Naast de constitutieve route zijn andere eiwitten bestemd voor de buitenkant van de cel en ontstaan door signalen uit de omgeving, of het nu gaat om hormonen, enzymen of neurotransmitters..
In pancreascellen worden spijsverteringsenzymen bijvoorbeeld verpakt in blaasjes die alleen worden uitgescheiden als de aanwezigheid van voedsel wordt gedetecteerd..
Recent onderzoek meldt het bestaan van alternatieve routes voor membraaneiwitten die niet door het Golgi-apparaat gaan. Deze paden omzeilen "Onconventioneel" wordt in de literatuur besproken.
Er zijn vijf modellen om de eiwithandel in het apparaat te verklaren. De eerste betreft het verkeer van materiaal tussen stalcompartimenten, elk heeft de nodige enzymen om specifieke functies te vervullen. Het tweede model omvat de geleidelijke rijping van de reservoirs.
De derde stelt ook de rijping van de zakken voor, maar met de opname van een nieuw onderdeel: het buisvormige transport. Volgens het model zijn tubuli belangrijk in het verkeer in beide richtingen.
Het vierde model stelt voor dat het complex als een eenheid werkt. Het vijfde en laatste model is het meest recente en stelt dat het complex is opgedeeld in verschillende compartimenten.
In bepaalde celtypen heeft het Golgi-complex specifieke functies. De cellen van de alvleesklier hebben gespecialiseerde structuren voor de afscheiding van insuline.
De verschillende bloedgroepen bij mensen zijn een voorbeeld van differentiële glycosylatiepatronen. Dit fenomeen wordt verklaard door de aanwezigheid van verschillende allelen die coderen voor glucotransferase.



Niemand heeft nog op dit artikel gereageerd.