De chromatine Het is het complex gevormd door DNA en eiwitten, uniek in eukaryote organismen. Qua verhoudingen bevat het bijna twee keer zoveel eiwitten als genetisch materiaal. De belangrijkste eiwitten in dit complex zijn histonen: kleine, positief geladen eiwitten die via elektrostatische interacties aan DNA binden. Bovendien heeft chromatine meer dan duizend andere eiwitten dan histonen..
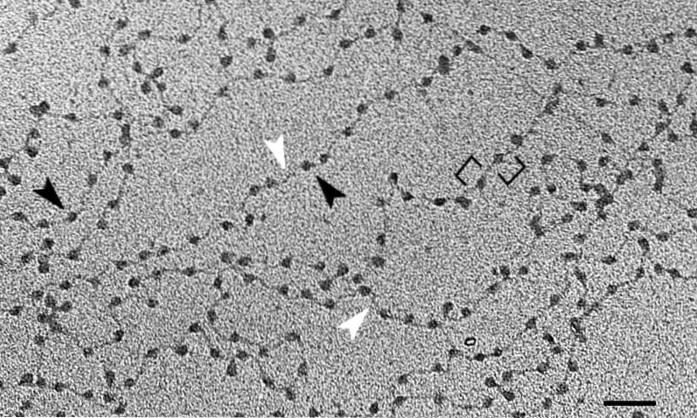
De fundamentele eenheid van chromatine is het nucleosoom, dat bestaat uit de vereniging van histonen en DNA. Dit arrangement doet denken aan de kralen aan een ketting. Nadat we alle hogere niveaus van DNA-organisatie hebben doorlopen, zullen we de chromosomen bereiken.
Chromatinestructuur is nauw verwant aan de controle van genexpressie. Er zijn twee hoofdtypen: euchromatine en heterochromatine.
Euchromatin wordt gekenmerkt door een lage verdichtingsgraad, wat zich vertaalt in hoge transcriptieniveaus. Heterochromatine daarentegen is transcriptioneel inactief vanwege de hoge mate van verdichting..
Structureel zijn er bepaalde epigenetische kenmerken van de histonen die kenmerkend zijn voor beide typen chromatinen. Hoewel euchromatine wordt geassocieerd met acetylering, wordt heterochromatine geassocieerd met een afname van chemische groepen.
Er zijn enkele regio's van heterochromatine met structurele functies, zoals telomeren en centromeren.
Artikel index
De studie van genetisch materiaal en de structurele organisatie ervan begon in 1831 toen de onderzoeker Robert Brown de kern beschreef. Een van de directe vragen bij deze ontdekking was om te informeren naar de biologische en chemische aard van deze structuur..
Deze vragen werden tussen 1870 en 1900 opgehelderd met de experimenten van Friedrich Miescher, die het woord nucleïne introduceerde. Walther Flemming wijzigt echter de term en gebruikt chromatine om te verwijzen naar de nucleaire substantie.
Met het verstrijken van de tijd begin je een dieper begrip te krijgen van het genetisch materiaal en zijn eigenschappen. Pas in 1908 merkte de Italiaanse onderzoeker Pasquale Baccarini dat het chromatine niet homogeen was en erin slaagde om kleine lichaampjes in de kern te visualiseren..
De typen chromatines - euchromatine en heterochromatine - werden aanvankelijk voorgesteld door Emil Heitz in 1928. Om deze classificatie vast te stellen, vertrouwde Heitz op het gebruik van kleurstoffen.
In 1974 stelde de bioloog Roger Kornberg een model voor voor de organisatie van genetisch materiaal in structuren die bekend staan als nucleosomen, een hypothese die empirisch werd bevestigd door de experimenten van Markus Noll..
Chromatine is een nucleoproteïne dat wordt gevormd door de vereniging van genetisch materiaal - DNA - met een heterogene reeks eiwitten. Deze associatie is zeer dynamisch en krijgt een complexe driedimensionale conformatie waardoor het zijn regulerende en structurele functies kan uitoefenen..
Een van de belangrijkste chromatine-eiwitten zijn histonen, die in bijna dezelfde verhouding voorkomen als DNA.
Histonen zijn basiseiwitten, opmerkelijk geconserveerd gedurende de evolutionaire geschiedenis van organische wezens - dat wil zeggen, onze histonen variëren niet veel in vergelijking met die van andere zoogdieren, zelfs een ander dier dat fylogenetisch verder verwijderd is..
De lading van histonen is positief, dus ze kunnen via elektrostatische krachten interageren met de negatieve lading van de fosfaatruggengraat die in DNA aanwezig is. Er zijn vijf soorten histonen, namelijk: H1, H2A, H2B, H3 en H4.
Er zijn ook een aantal eiwitten van een andere aard dan histonen die deelnemen aan DNA-verdichting..
De basiseenheid van chromatine zijn nucleosomen - repetitieve structuren die bestaan uit DNA en histonen, een conformatie die overal in genetisch materiaal voorkomt..
De dubbele DNA-helix is gewikkeld in een complex van acht histonen die bekend staan als histone octamer. Het molecuul rolt in ongeveer twee windingen, gevolgd door een kort gebied (tussen 20 en 60 basenparen) dat de nucleosomen van elkaar scheidt..
Om deze organisatie te begrijpen, moeten we er rekening mee houden dat het DNA-molecuul extreem lang is (ongeveer 2 meter) en op een ordelijke manier moet worden opgewonden om zich in de kern te vestigen (met een diameter van 3 tot 10 µm). Bovendien moet het beschikbaar worden gesteld voor replicatie en transcriptie..
Dit doel wordt bereikt met verschillende niveaus van DNA-verdichting, de eerste zijn de bovengenoemde nucleosomen. Deze lijken op de kralen aan een parelketting. Ongeveer 150 basenparen van DNA worden thuis 'geteld' gewikkeld.
Er zijn geen waargebeurde verhalen over bacteriën. Daarentegen is er een reeks eiwitten die op histonen lijken en er wordt aangenomen dat deze bijdragen aan de verpakking van bacterieel DNA..
De organisatie van chromatine is niet beperkt op het nucleosoomniveau. Deze associatie van eiwitten en DNA is gegroepeerd in een dikkere structuur van ongeveer 30 nm - vanwege deze dikte wordt het het "30 nm vezel" -niveau genoemd..
Het chromatine georganiseerd in de dikte van 30 nm is op zijn beurt georganiseerd in de vorm van lussen die zich uitstrekken in een soort steiger van eiwitachtige aard (geen histonen).
Dit is het model dat momenteel wordt gebruikt, hoewel het bestaan van complexere verdichtingsmechanismen kan worden verwacht. De uiteindelijke organisatie bestaat uit het chromosoom.
De verdichting en organisatie van genetisch materiaal is van vitaal belang voor meerdere biologische functies. Verschillende medische aandoeningen zijn in verband gebracht met fouten in de chromatinestructuur, waaronder onder meer X-gebonden alfa-thalassemie, het Rubinstein-Taybi-syndroom, het Coffin-Lowry-syndroom en het Rett-syndroom..
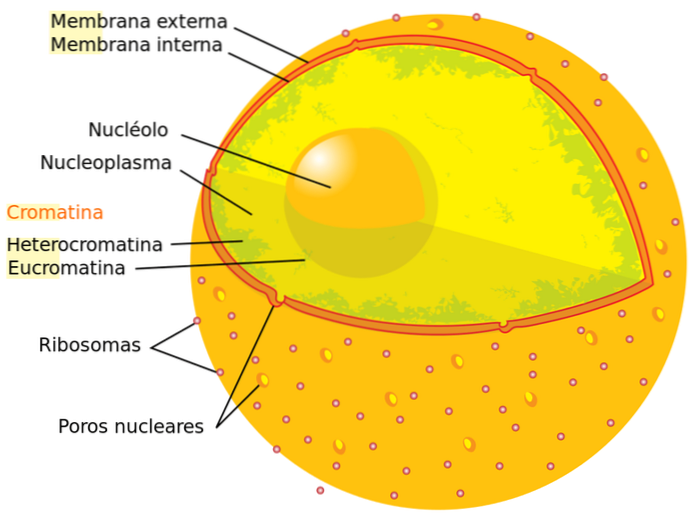
Er zijn twee soorten chromatine in de cel, die worden onthuld door het aanbrengen van kleurstoffen: euchromatine ("echt" chromatine) en heterochromatine. In het eerste geval wordt de kleuring zwak waargenomen, terwijl in het tweede geval de kleuring intens is.
Deze structurele organisatie van DNA is uniek voor eukaryote organismen en is cruciaal voor het gedrag van chromosomen en de regulatie van genexpressie..
Als we de verhoudingen van beide typen chromatines evalueren in een cel die zich in interfase bevindt, zien we dat ongeveer 90% van het chromatine euchromatine is en de resterende 10% overeenkomt met heterochromatine. We zullen elk type hieronder in detail beschrijven:
Het belangrijkste verschil tussen beide typen chromatine heeft te maken met de mate van verdichting of "pakking" van het molecuul tijdens specifieke stadia van celdeling..
Hoewel op het grensvlak het genetisch materiaal willekeurig verspreid lijkt te zijn, is het niet op deze manier.
Er is in dit stadium een aanzienlijke organisatie, waarbij een differentiële verdeling van het chromosomale materiaal te zien is in de kern.
Het DNA van chromosomen verstrengelt zich niet met de DNA-streng van andere chromosomen en blijft in specifieke regio's die chromosomale territoria worden genoemd. Deze organisatie lijkt bij te dragen aan genexpressie.
Heterochromatine is sterk gecondenseerd, waardoor het ontoegankelijk is voor de transcriptiemachines - dus het wordt niet getranscribeerd. Bovendien is het slecht in termen van het aantal genen dat het heeft..
Bepaalde regio's van heterochromatine zijn persistent in alle cellijnen - d.w.z., voor altijd het zal zich gedragen als heterochromatine. Dit type heterochromatine staat bekend als constitutief. Een voorbeeld hiervan zijn de gecondenseerde gebieden van chromosomen die centromeren en telomeren worden genoemd..
Daarentegen zijn er delen van heterochromatine die het verdichtingsniveau kunnen variëren als reactie op veranderingen in ontwikkelingspatronen of omgevingsvariabelen..
Dankzij nieuwe studies wordt deze visie opnieuw geformuleerd en nu zijn er aanwijzingen dat constitutief heterochromatine ook dynamisch is en in staat is om op prikkels te reageren..
Een van de factoren die de structuur van chromatine bepalen, zijn de chemische modificaties van histonen. In het geval van chromatine dat transcriptioneel inactief is, vertonen ze gehypoacetyleerde histonen.
De afname van het aantal acetylgroepen is geassocieerd met genuitschakeling, omdat het de positieve lading van lysines niet maskeert, waardoor een sterke elektrostatische interactie tussen DNA en histonen mogelijk is..
Een ander epigenetisch merk is methylering. Aangezien de toevoeging van een methylgroep de lading van het eiwit echter niet wijzigt, is de consequentie ervan (het activeren of deactiveren van genen) niet zo duidelijk en zal het afhangen van het gebied van de histon waar het merkteken wordt gevonden..
Empirisch is gevonden dat de methylering van H3K4me3 en H3K36me3 geassocieerd is met genactivering en die van H3K9me3 en H3K27me3..
In het voorbeeld van constitutief heterochromatine noemen we het centromeer. Dit chromosomale gebied speelt een structurele rol en draagt bij aan de beweging van chromosomen tijdens zowel mitotische als meiotische celdelingsgebeurtenissen..
In tegenstelling tot heterochromatine is euchromatine een minder compact molecuul, dus de transcriptiemachine heeft gemakkelijke toegang (met name het RNA-polymerase-enzym) en kan actief tot expressie worden gebracht via genetische routes.
De nucleosoomstructuur van een transcriptioneel actief chromatine wordt gekenmerkt door geacetyleerde histonen. De aanwezigheid van monomethyleerde lysines wordt ook geassocieerd met genactivering.
De toevoeging van een acetylgroep aan deze histonlysineresten neutraliseert de positieve lading van genoemd aminozuur. Het directe gevolg van deze verandering is de vermindering van elektrostatische interacties tussen histon en DNA, waardoor een losser chromatine ontstaat..
Deze structurele wijziging maakt de interacties mogelijk van het genetisch materiaal met de transcriptionele machinerie, die wordt gekenmerkt door bijzonder omvangrijk te zijn..
Euchromatin omvat alle genen die actief zijn en kunnen worden bereikt door de enzymatische machinerie die verband houdt met transcriptie. Daarom zijn de functies net zo breed als de functies van de betrokken genen..



Niemand heeft nog op dit artikel gereageerd.