
De sfingolipiden ze vertegenwoordigen een van de drie belangrijkste families van lipiden die aanwezig zijn in biologische membranen. Net als glycerofosfolipiden en sterolen zijn het amfipatische moleculen met een hydrofiel polair gebied en een hydrofoob apolair gebied..
Ze werden voor het eerst beschreven in 1884 door Johann L. W. Thudichum, die drie sfingolipiden (sfingomyeline, cerebrosiden en cerebrosulfatide) beschreef die tot de drie verschillende bekende klassen behoren: fosfosfingolipiden, neutrale en zure glycosfingolipiden..
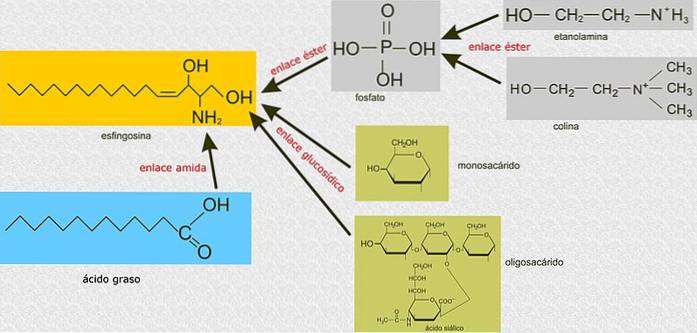
In tegenstelling tot glycerofosfolipiden, zijn sfingolipiden niet gebouwd op een glycerol-3-fosfaatmolecuul als het hoofdskelet, maar zijn het verbindingen die zijn afgeleid van sfingosine, een aminoalcohol met een lange koolwaterstofketen verbonden door een amidebinding..
In termen van complexiteit en diversiteit zijn er minstens 5 verschillende basistypen bekend voor sfingolipiden bij zoogdieren. Deze basen kunnen worden verbonden door meer dan 20 verschillende soorten vetzuren, met verschillende lengtes en verzadigingsgraden, naast de vele variaties in de polaire groepen die kunnen voorkomen..
Biologische membranen hebben ongeveer 20% sfingolipiden. Deze hebben verschillende en belangrijke functies in cellen, van structurele tot signaaltransductie en controle van verschillende cellulaire communicatieprocessen..
De verdeling van deze moleculen varieert afhankelijk van de functie van het organel waar ze worden aangetroffen, maar normaal gesproken is de concentratie van sfingolipiden veel hoger in de buitenste monolaag van het plasmamembraan in vergelijking met de binnenste monolaag en andere compartimenten..
Bij mensen zijn er minstens 60 soorten sfingolipiden. Velen van hen zijn belangrijke componenten van de membranen van zenuwcellen, terwijl andere een belangrijke structurele rol spelen of deelnemen aan onder meer signaaltransductie, herkenning, celdifferentiatie, pathogenese, geprogrammeerde celdood..
Artikel index
Alle sfingolipiden zijn afgeleid van een L-serine, die wordt gecondenseerd met een vetzuur met een lange keten om de sfingoïde basis te vormen, ook wel bekend als de lange keten base (LCB)..
De meest voorkomende basen zijn sfinganine en sfingosine, die alleen van elkaar verschillen in de aanwezigheid van een dubbele binding trans tussen koolstofatomen 4 en 5 van het vetzuur van sfingosine.
De koolstofatomen 1, 2 en 3 van sfingosine zijn structureel analoog aan de koolstofatomen van glycerol van glycerofosfolipiden. Wanneer een vetzuur via amidebindingen aan koolstof 2 van sfingosine wordt gehecht, wordt een ceramide geproduceerd, een molecuul dat sterk lijkt op diacylglycerol en het eenvoudigste sfingolipide vertegenwoordigt..
De vetzuren met lange keten waaruit de hydrofobe gebieden van deze lipiden bestaan, kunnen zeer divers zijn. De lengtes variëren van 14 tot 22 koolstofatomen die verschillende verzadigingsgraden kunnen hebben, meestal tussen koolstofatomen 4 en 5.
Op posities 4 of 6 kunnen ze hydroxylgroepen hebben en dubbele bindingen op andere posities of zelfs vertakkingen zoals methylgroepen.
De vetzuurketens die door amidebindingen aan ceramiden zijn verbonden, zijn gewoonlijk verzadigd en hebben de neiging langer te zijn dan die in glycerofosfolipiden, wat cruciaal lijkt te zijn voor hun biologische activiteit..
Een onderscheidend kenmerk van het sfingolipideskelet is dat ze een netto positieve lading kunnen hebben bij neutrale pH, wat zeldzaam is bij lipidemoleculen..
Echter, de pKa van de aminogroep is laag in vergelijking met een eenvoudig amine, tussen 7 en 8, zodat een deel van het molecuul niet wordt geladen bij fysiologische pH, wat de "vrije" beweging hiervan tussen de dubbellagen zou kunnen verklaren.
De traditionele classificatie van sfingolipiden komt voort uit de veelvoudige modificaties die het ceramidemolecuul kan ondergaan, vooral wat betreft de substituties van de groepen van de polaire kop..
Sfingolipiden zijn essentieel bij dieren, planten en schimmels, evenals bij sommige prokaryote organismen en virussen..
Sfingolipiden moduleren de fysische eigenschappen van membranen, inclusief hun vloeibaarheid, dikte en kromming. Door deze eigenschappen te moduleren, hebben ze ook directe invloed op de ruimtelijke organisatie van membraaneiwitten..
In biologische membranen kunnen dynamische microdomeinen met minder vloeibaarheid worden gedetecteerd, bestaande uit cholesterol- en sfingolipidemoleculen die lipidevlotten worden genoemd..
Deze structuren komen van nature voor en zijn nauw verwant aan integrale eiwitten, celoppervlakreceptoren en signaaleiwitten, transporteiwitten en andere eiwitten met glycosylfosfatidylinositol (GPI) -ankers..
Ze hebben functies als signaalmoleculen die fungeren als tweede boodschappers of als uitgescheiden liganden voor receptoren op het celoppervlak..
Als secundaire boodschappers kunnen ze deelnemen aan de regulatie van calciumhomeostase, celgroei, tumorvorming en onderdrukking van apoptose. Bovendien hangt de activiteit van veel integrale en perifere membraaneiwitten af van hun associatie met sfingolipiden..
Veel interacties tussen cellen en de cel met zijn omgeving zijn afhankelijk van de blootstelling van de verschillende polaire groepen sfingolipiden aan de buitenkant van het plasmamembraan..
De binding van glycosfingolipiden en lectines is cruciaal voor de associatie van myeline met axonen, de adhesie van neutrofielen aan het endotheel, enz..
De belangrijkste signalerende sfingolipiden zijn basen met lange ketens of sfingosinen en ceramiden, evenals hun gefosforyleerde derivaten, zoals sfingosine-1-fosfaat.
De metabole producten van veel sfingolipiden activeren of remmen meerdere stroomafwaartse doelwitten (proteïnekinasen, fosfoproteïnefosfatasen en andere), die complexe cellulaire gedragingen regelen, zoals groei, differentiatie en apoptose..
Sommige pathogenen gebruiken glycosfingolipiden als receptoren om hun intrede in gastheercellen te bemiddelen of om virulentiefactoren aan hen af te leveren..
Van sfingolipiden is aangetoond dat ze deelnemen aan meerdere cellulaire gebeurtenissen zoals secretie, endocytose, chemotaxis, neurotransmissie, angiogenese en ontsteking.
Ze zijn ook betrokken bij membraantransport en beïnvloeden zo de internalisatie van receptoren, de ordening, beweging en fusie van secretoire blaasjes als reactie op verschillende stimuli..
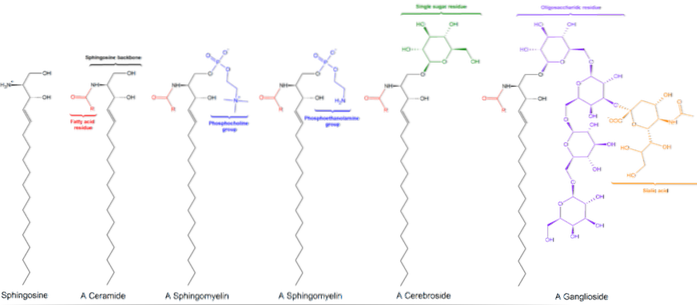
Er zijn drie subklassen van sfingolipiden, alle afgeleid van ceramide en verschillen van elkaar door polaire groepen, namelijk: sfingomyelines, glycolipiden en gangliosiden..

Deze bevatten fosfocholine of fosfoethanolamine als een polaire kopgroep, daarom worden ze samen met glycerofosfolipiden geclassificeerd als fosfolipiden. Ze lijken natuurlijk op fosfatidylcholines in driedimensionale structuur en algemene eigenschappen, aangezien ze geen lading hebben op hun polaire koppen..
Ze zijn aanwezig in de plasmamembranen van dierlijke cellen en komen vooral voor in myeline, een omhulsel dat de axonen van sommige neuronen omgeeft en isoleert..
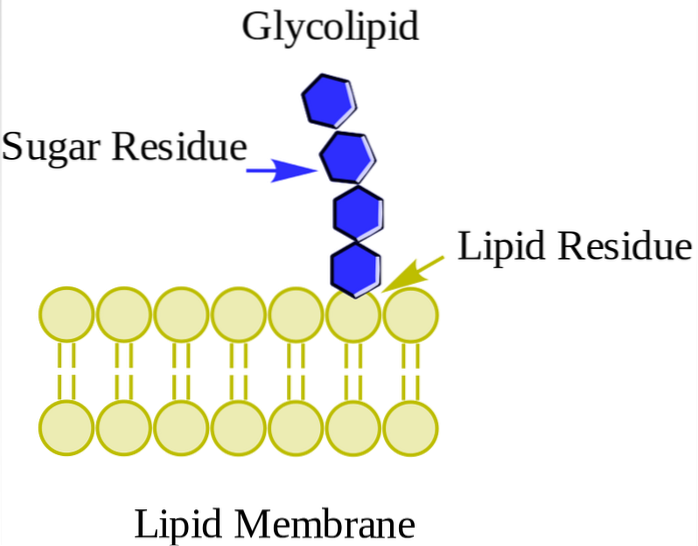
Ze worden voornamelijk aangetroffen op de buitenzijde van het plasmamembraan en hebben een of meer suikers als polaire kopgroep die rechtstreeks is bevestigd aan de hydroxylgroep van koolstof 1 van het ceramidegedeelte. Ze hebben geen fosfaatgroepen. Omdat ze bij pH 7 geen lading hebben, worden ze neutrale glycolipiden genoemd.
Cerebrosides hebben een enkel suikermolecuul dat aan ceramide is gehecht. Degenen die galactose bevatten, worden aangetroffen in plasmamembranen van niet-zenuwweefselcellen. Globosiden zijn glycosfingolipiden met twee of meer suikers, meestal D-glucose, D-galactose of N-acetyl-D-galactosamine..
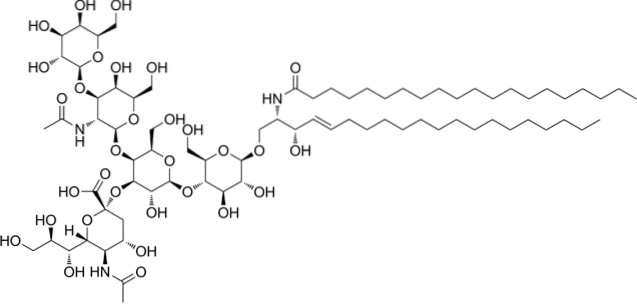
Dit zijn de meest complexe sfingolipiden. Ze hebben oligosacchariden als polaire kopgroep en een of meer terminale N-acetylmuraminezuurresiduen, ook wel siaalzuur genoemd. Siaalzuur geeft gangliosiden een negatieve lading bij pH 7, waardoor ze zich onderscheiden van neutrale glycosfingolipiden.
De nomenclatuur van deze klasse van sfingolipiden hangt af van de hoeveelheid siaalzuurresiduen die aanwezig zijn in het oligosaccharidegedeelte van de polaire kop..
Het basismolecuul met lange keten, of sfingosine, wordt gesynthetiseerd in het endoplasmatisch reticulum (ER) en de toevoeging van de polaire groep aan de kop van deze lipiden vindt later plaats in het Golgi-complex. Bij zoogdieren kan enige synthese van sfingolipiden ook plaatsvinden in de mitochondriën.
Na voltooiing van hun synthese in het Golgi-complex, worden sfingolipiden getransporteerd naar andere celcompartimenten via vesikel-gemedieerde mechanismen..
De biosynthese van sfingolipiden bestaat uit drie fundamentele gebeurtenissen: de synthese van basen met lange ketens, de biosynthese van ceramiden door de vereniging van een vetzuur door een amidebinding en tenslotte de vorming van complexe sfingolipiden door de vereniging van de polaire groepen. op koolstof 1 van de sfingoïde basis.
Naast de synthese de novo, sfingolipiden kunnen ook worden gevormd door de omzet of recycling van basen met lange ketens en ceramiden, die de pool van sfingolipiden kunnen voeden.
De biosynthese van ceramide, de ruggengraat van sfingolipiden, begint met de decarboxylatieve condensatie van een palmitoyl-CoA-molecuul en een L-serine. De reactie wordt gekatalyseerd door een heterodimere serinepalmitoyltransferase (SPT), afhankelijk van pyridoxaalfosfaat en het product is 3-keto-dihydrosfingosine..
Dit enzym wordt geremd door β-halo-L-alanines en L-cycloserines. In gist wordt het gecodeerd door twee genen, terwijl er bij zoogdieren drie genen voor dit enzym zijn. De actieve site bevindt zich op het cytoplasmatische oppervlak van het endoplasmatisch reticulum.
De rol van dit eerste enzym is behouden in alle bestudeerde organismen. Er zijn echter enkele verschillen tussen de taxa die te maken hebben met de subcellulaire locatie van het enzym: die van bacteriën is cytoplasmatisch, die van gisten, planten en dieren zit in het endoplasmatisch reticulum..
3-ketosphinganine wordt vervolgens gereduceerd door NADPH-afhankelijke 3-ketosphinganine reductase om sfinganine te produceren. Dihydroceramidesynthase (sfinganine N-acyltransferase) acetyleert vervolgens sfinganine om dihydroceramide te produceren. Ceramide wordt vervolgens gevormd door dihydroceramide desaturase / reductase, dat een dubbele binding invoegt trans in positie 4-5.
Bij zoogdieren zijn er talrijke isovormen van ceramidesynthasen, die elk een specifieke keten van vetzuren binden aan basen met een lange keten. Daarom vormen ceramidesynthases en andere enzymen, elongases, de belangrijkste bron van diversiteit voor vetzuren in sfingolipiden..
Sfingomyeline wordt gesynthetiseerd door de overdracht van een fosfocholine van fosfatidylcholine naar ceramide, waarbij diacylglycerol vrijkomt. De reactie bindt de signaalroutes van sfingolipiden en glycerofosfolipiden.
Fosfoethanolamine ceramide wordt gesynthetiseerd uit fosfatidylethanolamine en ceramide in een reactie analoog aan die van sfingomyeline synthese, en eenmaal gevormd kan het worden gemethyleerd tot sfingomyeline. Inositolfosfaatceramiden worden gevormd door omestering uit fosfatidylinositol.
Glycosfingolipiden worden voornamelijk gemodificeerd in het Golgi-complex, waar specifieke glycosyltransferase-enzymen deelnemen aan de toevoeging van oligosaccharideketens in het hydrofiele gebied van het ceramideskelet..
De afbraak van sfingolipiden wordt uitgevoerd door de enzymen glucohydrolasen en sfingomyelinasen, die verantwoordelijk zijn voor het verwijderen van de modificaties van de polaire groepen. Aan de andere kant regenereren ceramidasen basen met lange ketens uit ceramiden.
Gangliosiden worden afgebroken door een reeks lysosomale enzymen die de stapsgewijze eliminatie van suikereenheden katalyseren en uiteindelijk een ceramide produceren..
Een andere afbraakroute bestaat uit de internalisatie van sfingolipiden in endocytische blaasjes die worden teruggestuurd naar het plasmamembraan of getransporteerd naar de lysosomen waar ze worden afgebroken door specifieke zure hydrolasen..
Niet alle basen met een lange keten worden gerecycled, het endoplasmatisch reticulum heeft een route voor hun terminale afbraak. Dit degradatiemechanisme bestaat uit fosforylering in plaats van acylering van LCB's, waardoor signaalmoleculen ontstaan die oplosbare substraten kunnen zijn voor lyase-enzymen die LCB's-fosfaat knippen om acyloaldehyden en fosfoethanolamine te genereren..
Het metabolisme van deze lipiden wordt op verschillende niveaus gereguleerd, een daarvan is dat van de enzymen die verantwoordelijk zijn voor de synthese, hun posttranslationele modificaties en hun allostere mechanismen..
Sommige regulatiemechanismen zijn celspecifiek, hetzij om het moment van celontwikkeling waarin ze worden geproduceerd te regelen, hetzij als reactie op specifieke signalen..


Niemand heeft nog op dit artikel gereageerd.