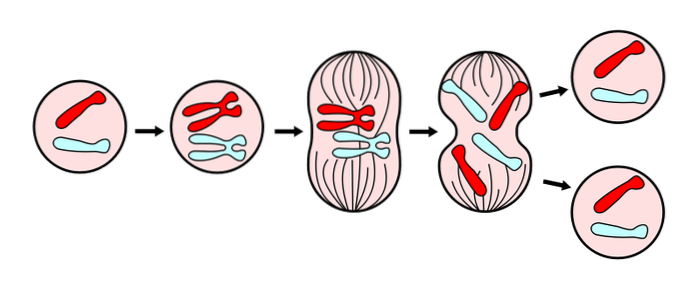
De mitose het is een proces van celdeling, waarbij een cel genetisch identieke dochtercellen produceert; voor elke cel worden twee "dochters" gegenereerd met dezelfde chromosomale belasting. Deze deling vindt plaats in de somatische cellen van eukaryote organismen..
Dit proces is een van de fasen van de celcyclus van eukaryote organismen, die bestaat uit 4 fasen: S (DNA-synthese), M (celdeling), G1 en G2 (tussenliggende fasen waarin mRNA's en eiwitten worden geproduceerd). Samen worden de G1-, G2- en S-fasen als een interface beschouwd. Nucleaire en cytoplasmatische deling (mitose en cytokinese) vormen de laatste fase van de celcyclus.
Op moleculair niveau wordt mitose geïnitieerd door de activering van een kinase (eiwit) genaamd MPF (Maturation Promoting Factor) en de daaruit voortvloeiende fosforylering van een aanzienlijk aantal van de samenstellende eiwitten van de cel. Met dit laatste kan de cel de morfologische veranderingen presenteren die nodig zijn om het delingsproces uit te voeren..
Mitose is een aseksueel proces, aangezien de stamcel en zijn dochters precies dezelfde genetische informatie hebben. Deze cellen staan bekend als diploïde omdat ze de volledige chromosomale belasting dragen (2n).
Meiose daarentegen is het proces van celdeling dat leidt tot seksuele voortplanting. In dit proces repliceert een diploïde stamcel zijn chromosomen en deelt hij zich vervolgens twee keer achter elkaar (zonder zijn genetische informatie te repliceren). Ten slotte worden 4 dochtercellen gegenereerd met slechts de helft van de chromosomale belasting, die haploïde (n) worden genoemd.
Artikel index
Mitose in eencellige organismen produceert over het algemeen dochtercellen die sterk lijken op hun voorlopers. Daarentegen kan dit proces tijdens de ontwikkeling van meercellige wezens aanleiding geven tot twee cellen met een aantal verschillende kenmerken (ondanks dat ze genetisch identiek zijn)..
Door deze celdifferentiatie ontstaan de verschillende celtypen waaruit meercellige organismen bestaan..
Tijdens het leven van een organisme vindt de celcyclus continu plaats, waarbij voortdurend nieuwe cellen worden gevormd die op hun beurt groeien en zich voorbereiden om te delen door mitose.
Celgroei en celdeling worden gereguleerd door mechanismen, zoals apoptose (geprogrammeerde celdood), die het mogelijk maken een evenwicht te bewaren en overmatige weefselgroei te vermijden. Op deze manier wordt ervoor gezorgd dat de defecte cellen worden vervangen door nieuwe cellen, in overeenstemming met de eisen en behoeften van het lichaam..
Het vermogen om zich voort te planten is een van de belangrijkste kenmerken van alle organismen (van eencellig tot meercellig) en van de cellen waaruit het bestaat. Deze kwaliteit zorgt voor de continuïteit van uw genetische informatie..
Het begrijpen van de processen van mitose en meiose hebben een fundamentele rol gespeeld bij het begrijpen van de intrigerende cellulaire kenmerken van organismen. Bijvoorbeeld de eigenschap om het aantal chromosomen constant te houden van de ene cel naar de andere binnen een individu en tussen individuen van dezelfde soort.
Wanneer we een soort snee of wond op onze huid krijgen, zien we hoe de beschadigde huid zich binnen een paar dagen herstelt. Dit gebeurt dankzij het proces van mitose.
In het algemeen volgt mitose dezelfde opeenvolging van processen (fasen) in alle eukaryote cellen. In deze fasen treden veel morfologische veranderingen op in de cel. Onder hen de condensatie van chromosomen, breuk van het kernmembraan, scheiding van de cel van de extracellulaire matrix en andere cellen, en de deling van het cytoplasma..
In sommige gevallen worden nucleaire deling en cytoplasmatische deling als afzonderlijke fasen beschouwd (respectievelijk mitose en cytokinese).
Voor een betere studie en begrip van het proces zijn zes (6) fasen aangewezen, genaamd: profase, prometafase, metafase, anafase en telofase, waarbij cytokinese wordt beschouwd als een zesde fase, die zich begint te ontwikkelen tijdens anafase.
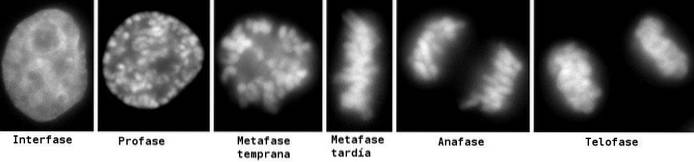
Deze fasen worden sinds de 19e eeuw door de lichtmicroscoop bestudeerd, dus tegenwoordig zijn ze gemakkelijk herkenbaar aan de morfologische kenmerken die de cel vertoont, zoals chromosomale condensatie en de vorming van de mitotische spoel..
Profase is de eerste zichtbare manifestatie van celdeling. In deze fase kan het verschijnen van chromosomen worden gezien als te onderscheiden vormen, vanwege de progressieve verdichting van chromatine. Deze condensatie van chromosomen begint met de fosforylering van Histone H1-moleculen door MPF-kinase..
Het condensatieproces bestaat uit het samentrekken en daarmee het verkleinen van de grootte van de chromosomen. Dit gebeurt door het oprollen van de chromatinevezels, waardoor gemakkelijker verplaatsbare structuren (mitotische chromosomen) ontstaan.
Chromosomen die eerder zijn gedupliceerd tijdens de S-periode van de celcyclus, krijgen een dubbelstrengs uiterlijk, zusterchromatiden genaamd, deze strengen worden bij elkaar gehouden door een gebied dat de centromeer wordt genoemd. In deze fase verdwijnen ook de nucleoli.
Tijdens de profase wordt de mitotische spoel gevormd, die bestaat uit microtubuli en eiwitten die een reeks vezels vormen.
Terwijl de spil zich vormt, worden de microtubuli van het cytoskelet gedemonteerd (door de eiwitten die hun structuur behouden, te deactiveren), waardoor het noodzakelijke materiaal wordt geleverd voor de vorming van de mitotische spil..
Het centrosoom (een membraanloos organel, functioneel in de celcyclus), gedupliceerd op het grensvlak, fungeert als de assemblage-eenheid van de spilmicrotubuli. In dierlijke cellen heeft het centrosoom een paar centriolen in het midden; maar deze zijn afwezig in de meeste plantencellen.
De gedupliceerde centrosomen beginnen van elkaar te scheiden terwijl de microtubuli van de spil in elk van hen worden geassembleerd en beginnen te migreren naar tegenoverliggende uiteinden van de cel..
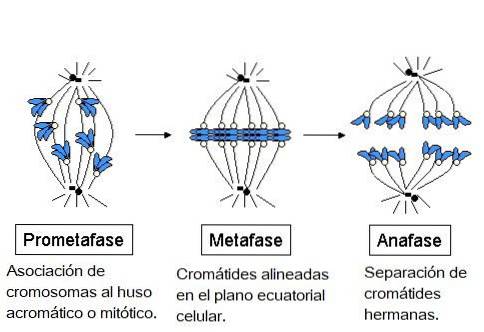
Aan het einde van de profase begint de breuk van de nucleaire envelop, die plaatsvindt in afzonderlijke processen: de demontage van de nucleaire porie, de nucleaire lamina en de nucleaire membranen. Door deze onderbreking kunnen de mitotische spil en chromosomen beginnen te interageren..
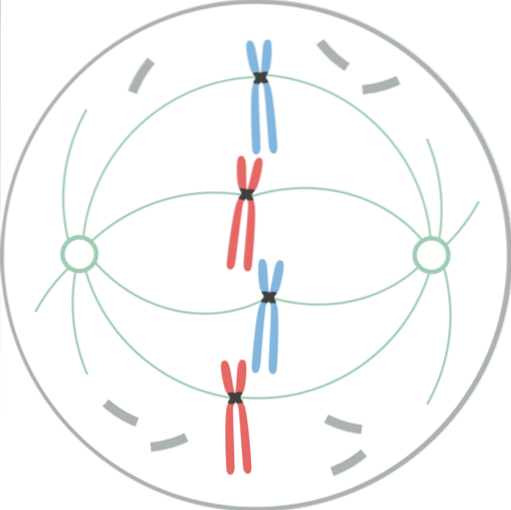
In dit stadium is de nucleaire envelop volledig gefragmenteerd, dus de microtubuli van de spil vallen dit gebied binnen en interageren met de chromosomen. De twee centrosomen zijn gescheiden, elk aan de polen van de mitotische spoel, aan tegenovergestelde uiteinden van de cellen..
Nu omvat de mitotische spoel de microtubuli (die zich uitstrekken van elk centrosoom naar het midden van de cel), de centrosomen en een paar asters (structuren met een radiale verdeling van korte microtubuli, die zich vanuit elk centrosoom ontvouwen)..
De chromatiden ontwikkelden elk een gespecialiseerde eiwitstructuur, de kinetochoor genaamd, die zich in het centromeer bevindt. Deze kinetochoor bevinden zich in tegengestelde richtingen en sommige microtubuli, microtubuli van de kinetochoor genaamd, hechten eraan..
Deze microtubuli, vastgemaakt aan de kinetochoor, beginnen naar het chromosoom te bewegen vanaf het einde waarvan ze zich uitstrekken; sommige van de ene pool en andere van de andere pool. Dit creëert een "trek- en krimp" -effect dat, wanneer gestabiliseerd, ervoor zorgt dat het chromosoom tussen de uiteinden van de cel terechtkomt..
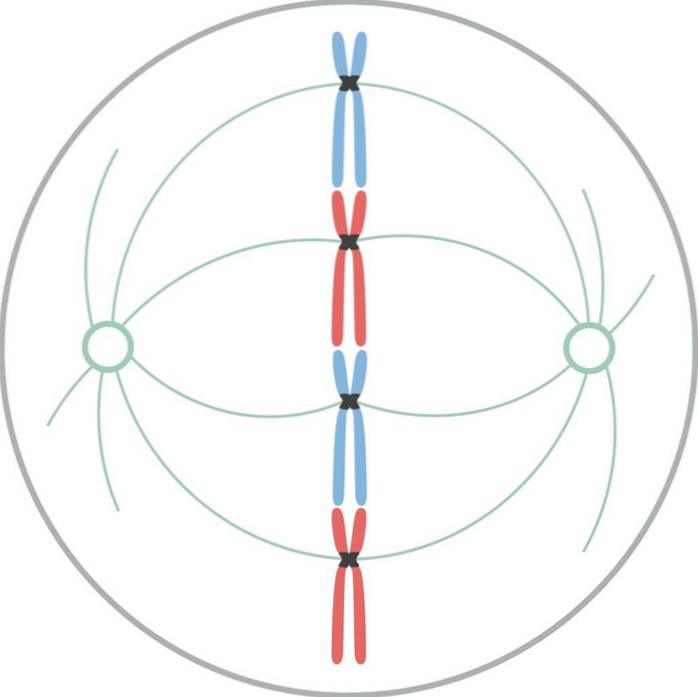
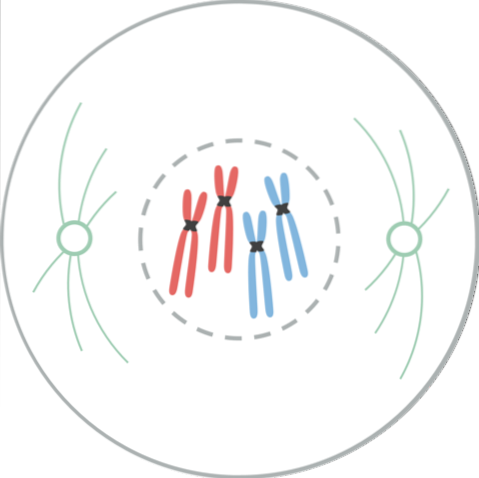
In metafase bevinden centrosomen zich aan tegenovergestelde uiteinden van cellen. De spil vertoont een duidelijke structuur, in het midden waarvan de chromosomen zich bevinden. De centromeren van deze chromosomen zijn aan de vezels vastgemaakt en uitgelijnd in een denkbeeldig vlak dat de metafaseplaat wordt genoemd..
De chromatidekinetochoren blijven gehecht aan de microtubuli van de kinetochoor. Microtubuli die niet aan de kinetochoren hechten en zich uitstrekken vanaf tegenovergestelde polen van de spil, werken nu met elkaar samen. Op dit punt zijn de microtubuli van de asters in contact met het plasmamembraan..
Deze groei en interactie van microtubuli completeert de structuur van de mitotische spoel en geeft het een "vogelkooi" -uiterlijk..
Morfologisch gezien is deze fase degene met de minste veranderingen, en daarom werd het beschouwd als een rustfase. Hoewel ze niet gemakkelijk merkbaar zijn, vinden er veel belangrijke processen in plaats, naast dat het de langste fase van mitose is..
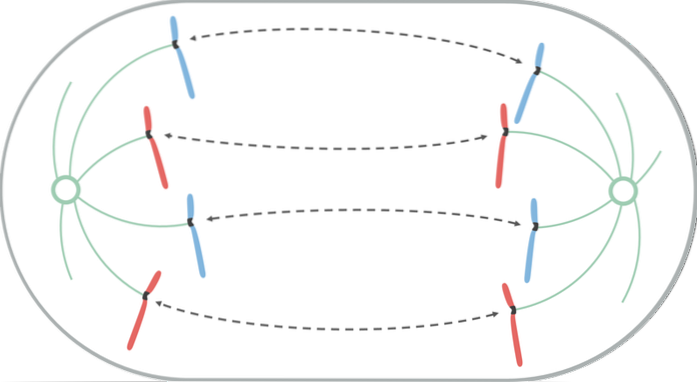
Tijdens anafase begint elk paar chromatiden te scheiden (vanwege de inactivering van de eiwitten die ze bij elkaar houden). Gescheiden chromosomen verplaatsen zich naar tegenovergestelde uiteinden van de cel.
Deze migratiebeweging is het gevolg van het korter worden van de microtubuli van de kinetochoor, waardoor een "trek" -effect ontstaat dat ervoor zorgt dat elk chromosoom van zijn centromeer beweegt. Afhankelijk van de locatie van het centromeer op het chromosoom, kan het tijdens zijn verplaatsing een bepaalde V- of J-vorm aannemen..
De microtubuli die niet aan de kinetochoor hechten, groeien en verlengen door de adhesie van tubuline (eiwit) en door de werking van motoreiwitten die eroverheen bewegen, waardoor het contact tussen hen stopt. Terwijl ze van elkaar af bewegen, doen de polen van de spil dat ook, waardoor de cel langer wordt.
Aan het einde van deze fase bevinden de groepen chromosomen zich aan tegenovergestelde uiteinden van de mitotische spoel, waardoor aan elk uiteinde van de cel een complete en gelijkwaardige set chromosomen overblijft..
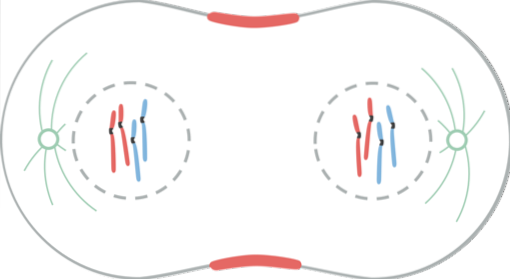
De telofase is de laatste fase van nucleaire deling. Kinetochore microtubuli vallen uiteen, terwijl polaire microtubuli verder verlengen.
Het kernmembraan begint zich rond elke set chromosomen te vormen, met behulp van de nucleaire enveloppen van de progenitorcel, die als blaasjes in het cytoplasma waren..
In dit stadium zijn de chromosomen die zich op de celpolen bevinden volledig gedecondenseerd door de defosforylering van de histonmoleculen (H1). De vorming van de elementen van het kernmembraan wordt gestuurd door verschillende mechanismen.
Tijdens anafase begonnen veel van de gefosforyleerde eiwitten in profase te defosforyleren. Hierdoor kunnen aan het begin van telofase de nucleaire blaasjes weer in elkaar worden gezet en associëren ze met het oppervlak van de chromosomen..
Aan de andere kant wordt de nucleaire porie weer in elkaar gezet, waardoor nucleaire eiwitten kunnen worden gepompt. De eiwitten van de nucleaire lamina worden gedefosforyleerd, waardoor ze weer kunnen associëren om de vorming van de nucleaire lamina te voltooien..
Ten slotte, nadat de chromosomen volledig zijn gedecondenseerd, wordt de RNA-synthese opnieuw gestart, waarbij de nucleoli opnieuw worden gevormd en zo de vorming van de nieuwe interfase-kernen van de dochtercellen wordt voltooid..

Cytokinese wordt beschouwd als een afzonderlijke gebeurtenis van nucleaire deling, en gewoonlijk begeleidt het cytoplasmatische delingsproces in typische cellen elke mitose, beginnend bij anafase. Verschillende onderzoeken hebben aangetoond dat bij sommige embryo's meerdere nucleaire delingen optreden voorafgaand aan cytoplasmatische deling.
Het proces begint met het verschijnen van een groef of spleet die is gemarkeerd in het vlak van de metafaseplaat, waardoor de scheiding plaatsvindt tussen de groepen chromosomen. De plaats van de spleet wordt specifiek aangegeven door de mitotische spoel, de microtubuli van de asters.
In de gemarkeerde spleet wordt een reeks microfilamenten aangetroffen die een ring vormen die naar de cytoplasmatische kant van het celmembraan is gericht en die grotendeels bestaat uit actine en myosine. Deze eiwitten interageren met elkaar waardoor de ring rond de groef kan samentrekken..
Deze contractie wordt gegenereerd door het glijden van de filamenten van deze eiwitten, wanneer ze met elkaar in wisselwerking staan, op dezelfde manier als bijvoorbeeld in spierweefsel..
De samentrekking van de annulus wordt dieper en oefent een "klemmend" effect uit dat uiteindelijk de voorlopercellen verdeelt, waardoor de dochtercellen met hun zich ontwikkelende cytoplasmatische inhoud kunnen worden gescheiden..
Plantencellen hebben een celwand, dus hun cytoplasmatische delingsproces is anders dan eerder beschreven en begint in telofase..
De vorming van een nieuwe celwand begint wanneer de microtubuli van de restspil worden geassembleerd, die de fragmoplast vormen. Deze cilindrische structuur bestaat uit twee sets microtubuli die aan hun uiteinden met elkaar zijn verbonden en waarvan de positieve polen zijn ingebed in een elektronisch bord in het equatoriale vlak..
Kleine blaasjes van het Golgi-apparaat, verpakt met celwandprecursoren, reizen door de microtubuli van de fragmoplast naar het equatoriale gebied en combineren om een celplaat te vormen. De inhoud van de blaasjes wordt uitgescheiden in deze plaat terwijl deze groeit.
Deze plaque groeit en versmelt met het plasmamembraan langs de celomtrek. Dit komt door de constante herschikking van de microtubuli van de fragmoplast aan de rand van de plaat, waardoor meer blaasjes naar dit vlak kunnen bewegen en hun inhoud kunnen legen..
Op deze manier vindt de cytoplasmatische scheiding van de dochtercellen plaats. Ten slotte zorgt de inhoud van de celplaat samen met de cellulosemicrovezels erin om de vorming van de nieuwe celwand te voltooien..
Mitose is een delingsmechanisme in cellen en maakt deel uit van een van de fasen van de celcyclus in eukaryoten. Op een eenvoudige manier kunnen we zeggen dat de belangrijkste functie van dit proces de reproductie van een cel in twee dochtercellen is.
Voor eencellige organismen betekent celdeling het genereren van nieuwe individuen, terwijl dit proces voor meercellige organismen deel uitmaakt van de groei en het correct functioneren van het hele organisme (celdeling genereert de ontwikkeling van weefsels en het onderhoud van structuren).
Het mitoseproces wordt geactiveerd volgens de vereisten van het organisme. Bij zoogdieren beginnen rode bloedcellen (erytrocyten) zich bijvoorbeeld te delen, waardoor er meer cellen worden gevormd, wanneer het lichaam een betere zuurstofopname nodig heeft. Evenzo planten witte bloedcellen (leukocyten) zich voort wanneer het nodig is om een infectie te bestrijden..
Daarentegen missen sommige gespecialiseerde dierlijke cellen praktisch het mitoseproces of is het erg traag. Voorbeelden hiervan zijn zenuwcellen en spiercellen).
Over het algemeen zijn het cellen die deel uitmaken van het bindweefsel en structurele weefsel van het lichaam en waarvan de voortplanting alleen nodig is als een cel een defect of verslechtering heeft en moet worden vervangen.
Het celdelings- en groeicontrolesysteem is veel complexer in meercellige organismen dan in eencellige organismen. In het laatste geval wordt de reproductie in wezen beperkt door de beschikbaarheid van bronnen.
In dierlijke cellen wordt de deling gestopt totdat er een positief signaal is dat dit proces activeert. Deze activering komt in de vorm van chemische signalen van naburige cellen. Dit maakt het mogelijk om de onbeperkte groei van weefsels en de reproductie van defecte cellen te voorkomen, wat het leven van het organisme ernstig kan schaden.
Een van de mechanismen die de celvermenigvuldiging regelen, is apoptose, waarbij een cel sterft (door de productie van bepaalde eiwitten die zelfvernietiging activeren) als deze aanzienlijke schade aanricht of geïnfecteerd is door een virus..
Er is ook de regulering van de celontwikkeling door het remmen van groeifactoren (zoals eiwitten). De cellen blijven dus op het grensvlak, zonder door te gaan naar de M-fase van de celcyclus..
Het mitoseproces vindt plaats in de overgrote meerderheid van eukaryote cellen, van eencellige organismen zoals gist, die het gebruiken als een aseksueel voortplantingsproces, tot complexe meercellige organismen zoals planten en dieren..
Hoewel de celcyclus in het algemeen hetzelfde is voor alle eukaryote cellen, zijn er opmerkelijke verschillen tussen eencellige en meercellige organismen. In het eerste geval wordt de groei en deling van cellen bevorderd door natuurlijke selectie. Bij meercellige organismen wordt de proliferatie beperkt door strikte controlemechanismen.
Bij eencellige organismen verloopt de voortplanting op een versnelde manier, aangezien de celcyclus constant werkt en de dochtercellen snel naar mitose gaan om door te gaan met deze cyclus. Terwijl de cellen van meercellige organismen aanzienlijk langer nodig hebben om te groeien en te delen.
Er zijn ook enkele verschillen tussen de mitotische processen van planten- en dierencellen, aangezien in sommige fasen van dit proces het mechanisme in principe echter op dezelfde manier werkt bij deze organismen..
Prokaryote cellen groeien en delen in het algemeen sneller dan eukaryote cellen.
Organismen met prokaryote cellen (over het algemeen eencellig of in sommige gevallen meercellig) missen een kernmembraan dat het genetisch materiaal in een kern isoleert, dus het is verspreid in de cel, in een gebied dat de nucleoïde wordt genoemd. Deze cellen hebben een circulair hoofdchromosoom.
De celdeling in deze organismen is dus veel directer dan in eukaryote cellen, zonder het beschreven mechanisme (mitose). In hen wordt reproductie uitgevoerd door een proces dat binaire splitsing wordt genoemd, waarbij DNA-replicatie begint op een specifieke plaats op het circulaire chromosoom (oorsprong van replicatie of OriC)..
Er worden dan twee oorsprong gevormd die naar tegenovergestelde zijden van de cel migreren terwijl replicatie plaatsvindt, en de cel strekt zich uit tot twee keer zo groot. Aan het einde van de replicatie groeit het celmembraan uit tot het cytoplasma en verdeelt het de voorlopercellen in twee dochters met hetzelfde genetische materiaal..
De evolutie van eukaryote cellen bracht een toename van de complexiteit van het genoom met zich mee. Dit betrof de ontwikkeling van meer uitgebreide verdelingsmechanismen.
Er zijn hypotheses die suggereren dat bacteriële deling het voorlopermechanisme van mitose is. Er is een bepaalde relatie gevonden tussen eiwitten die geassocieerd zijn met binaire splitsing (dat kunnen eiwitten zijn die chromosomen verankeren aan specifieke plaatsen op het plasmamembraan van dochters) met tubuline en actine in eukaryote cellen..
Sommige studies wijzen op bepaalde eigenaardigheden bij de verdeling van moderne eencellige protisten. Bij hen blijft het kernmembraan intact tijdens mitose. De gerepliceerde chromosomen blijven verankerd op bepaalde plaatsen op dit membraan en scheiden zich wanneer de kern begint uit te rekken tijdens de celdeling..
Dit toont enig toeval aan met het proces van binaire splitsing, waarbij de gerepliceerde chromosomen zich hechten aan bepaalde plaatsen op het celmembraan. De hypothese stelt vervolgens voor dat de protisten die deze kwaliteit tijdens hun celdeling presenteren, deze eigenschap van een voorouderlijke prokaryote cel hadden kunnen behouden..
Op dit moment zijn er nog geen verklaringen ontwikkeld waarom het in eukaryote cellen van meercellige organismen nodig is dat het kernmembraan uiteenvalt tijdens het proces van celdeling..



Niemand heeft nog op dit artikel gereageerd.