
De celkern het is een fundamenteel compartiment van eukaryote cellen. Het is de meest opvallende structuur van dit celtype en heeft het erfelijk materiaal. Het stuurt alle cellulaire processen aan: het bevat alle instructies die in het DNA zijn gecodeerd om de nodige reacties uit te voeren. Het is betrokken bij de celdelingsprocessen.
Alle eukaryote cellen hebben een kern, behalve enkele specifieke voorbeelden zoals volwassen rode bloedcellen (erytrocyten) bij zoogdieren en floëemcellen in planten. Evenzo zijn er cellen met meer dan één kern, zoals sommige spiercellen, hepatocyten en neuronen..
De kern werd in 1802 ontdekt door Franz Bauer; In 1830 observeerde de wetenschapper Robert Brown echter ook deze structuur en werd hij populair als de belangrijkste ontdekker ervan. Door zijn grote formaat kan hij duidelijk onder een microscoop worden waargenomen. Bovendien is het een structuur die gemakkelijk te beitsen is..
De kern is geen homogene en statische bolvormige entiteit met verspreid DNA. Het is een complexe en ingewikkelde structuur met verschillende componenten en onderdelen erin. Bovendien is het dynamisch en verandert het voortdurend gedurende de celcyclus..
Artikel index
De kern is de hoofdstructuur die het mogelijk maakt onderscheid te maken tussen eukaryote en prokaryote cellen. Het is het grootste celcompartiment. Over het algemeen bevindt de kern zich dicht bij het midden van de cel, maar er zijn uitzonderingen, zoals plasmacellen en epitheelcellen..
Het is een bolvormig organel met een gemiddelde diameter van ongeveer 5 µm, maar kan, afhankelijk van het type cel, 12 µm bereiken. Ik kan ongeveer 10% van het totale celvolume innemen.
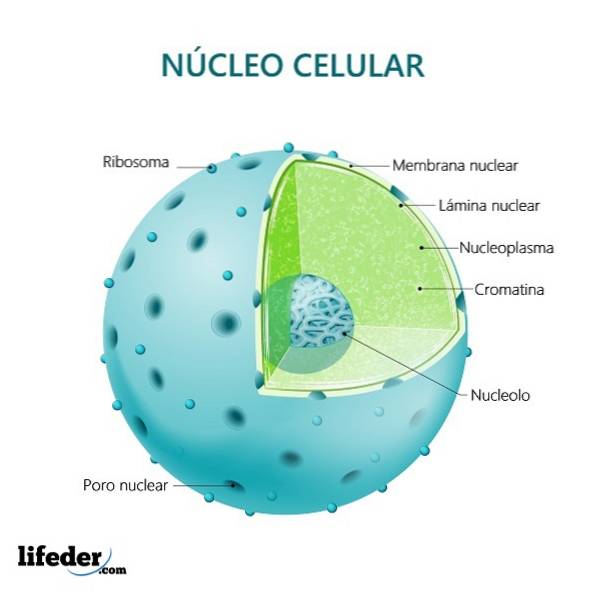
Het heeft een nucleaire envelop gevormd door twee membranen die het van het cytoplasma scheiden. Genetisch materiaal is samen met eiwitten erin georganiseerd.
Ondanks het feit dat er geen andere vliezige subcompartimenten in de kern zijn, kan een reeks componenten of gebieden binnen de structuur worden onderscheiden die specifieke functies hebben..
De kern krijgt een buitengewoon aantal functies toegewezen, omdat het de verzameling van alle genetische informatie van de cel bevat (met uitzondering van mitochondriaal DNA en chloroplast-DNA) en de processen van celdeling aanstuurt. Samengevat zijn de belangrijkste functies van de kernel als volgt:
Het bestaan van een lipidenbarrière tussen het genetisch materiaal en de rest van de cytoplasmatische componenten helpt de interferentie van andere componenten in het functioneren van DNA te verminderen. Dit vertegenwoordigt een evolutionaire innovatie die van groot belang is voor groepen eukaryoten..
Het splitsingsproces van boodschapper-RNA vindt plaats in de kern, voordat het molecuul naar het cytoplasma reist.
Het doel van dit proces is het elimineren van introns (“stukjes” genetisch materiaal die niet coderen en die de exons onderbreken, gebieden die coderen) uit het RNA. Vervolgens verlaat het RNA de celkern, waar het wordt vertaald in eiwitten..
Er zijn andere, meer specifieke functies van elke kernstructuur die later zullen worden besproken..
De kern bestaat uit drie gedefinieerde delen: de nucleaire envelop, de chromatine en de nucleolus. We zullen elke structuur hieronder in detail beschrijven:
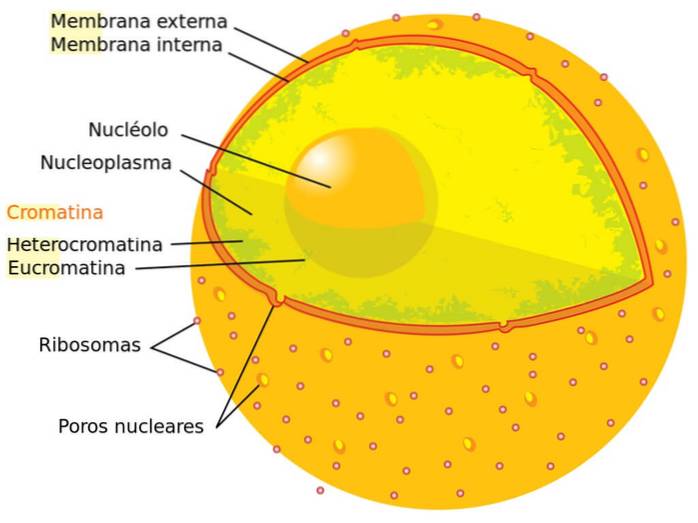
De nucleaire envelop is samengesteld uit membranen van lipide aard en scheidt de kern van de rest van de cellulaire componenten. Dit membraan is dubbel en daartussen bevindt zich een kleine ruimte die de perinucleaire ruimte wordt genoemd.
Het binnenste en buitenste membraansysteem vormt een continue structuur met het endoplasmatisch reticulum
Dit membraansysteem wordt onderbroken door een reeks poriën. Deze nucleaire kanalen maken de uitwisseling van materiaal met het cytoplasma mogelijk omdat de kern niet volledig geïsoleerd is van de rest van de componenten..
Door deze poriën vindt de uitwisseling van stoffen op twee manieren plaats: passief, zonder dat er energieverbruik nodig is; of actief, met energieverbruik. Passief kunnen kleine moleculen zoals water of zouten, kleiner dan 9 nm of 30-40 kDa, binnenkomen en vertrekken.
Dit gebeurt in tegenstelling tot moleculen met een hoog molecuulgewicht, die ATP (energie-adenosinetrifosfaat) nodig hebben om door deze compartimenten te bewegen. Grote moleculen zijn onder meer stukjes RNA (ribonucleïnezuur) of andere biomoleculen van eiwitachtige aard.
Poriën zijn niet zomaar gaten waar moleculen doorheen gaan. Het zijn grote eiwitstructuren, die 100 of 200 eiwitten kunnen bevatten en die "kernporiëncomplex" worden genoemd. Structureel lijkt het veel op een basketbalring. Deze eiwitten worden nucleoporines genoemd..
Dit complex is in een groot aantal organismen aangetroffen: van gisten tot mensen. Naast de cellulaire transportfunctie is het ook betrokken bij de regulatie van genexpressie. Ze zijn een onmisbare structuur voor eukaryoten.
Qua grootte en aantal kan het complex een grootte bereiken van 125 MDa bij gewervelde dieren, en een kern in deze diergroep kan ongeveer 2000 poriën hebben. Deze kenmerken variëren naargelang het bestudeerde taxon..
Chromatine wordt in de kern aangetroffen, maar we kunnen het niet als een compartiment ervan beschouwen. Het krijgt deze naam vanwege het uitstekende vermogen om te worden gekleurd en door de microscoop bekeken te worden.
DNA is een extreem lang lineair molecuul in eukaryoten. De verdichting is een sleutelproces. Het genetisch materiaal wordt geassocieerd met een reeks eiwitten, histonen genaamd, die een hoge affiniteit hebben voor DNA. Er zijn ook andere soorten eiwitten die kunnen interageren met DNA en geen histonen zijn.
In histonen spoelen DNA zich op en vormen chromosomen. Dit zijn dynamische structuren en hebben niet constant hun typische vorm (de X'en en Y'en die we gewend zijn te zien in boekillustraties). Deze regeling komt alleen voor tijdens de celdelingsprocessen.
In de rest van de stadia (wanneer de cel niet bezig is met delen), kunnen de individuele chromosomen niet worden onderscheiden. Dit feit suggereert niet dat de chromosomen homogeen of wanordelijk door de kern zijn verspreid..
Op de interface zijn de chromosomen georganiseerd in specifieke domeinen. In zoogdiercellen bezet elk chromosoom een specifiek 'territorium'.
Er kunnen twee soorten chromatine worden onderscheiden: heterochromatine en euchromatine. De eerste is sterk gecondenseerd en bevindt zich aan de rand van de kern, dus de transcriptiemachine heeft geen toegang tot deze genen. Euchromatin is losser georganiseerd.
Heterochromatine is onderverdeeld in twee soorten: constitutief heterochromatine, dat nooit tot expressie komt; en facultatief heterochromatine, dat in sommige cellen niet wordt getranscribeerd en in andere wel.
Het bekendste voorbeeld van heterochromatine als regulator van genexpressie is de condensatie en inactivering van het chromosoom X. Bij zoogdieren hebben vrouwtjes XX geslachtschromosomen, terwijl mannetjes XY zijn..
Om redenen van gendosering kunnen vrouwtjes niet tweemaal zoveel genen in X hebben als mannetjes. Om dit conflict te vermijden, wordt in elke cel een X-chromosoom willekeurig geïnactiveerd (en heterochromatine)..
De nucleolus is een zeer relevante interne structuur van de kern. Het is geen compartiment dat wordt afgebakend door vliezige structuren, het is een donkerder deel van de kern met specifieke functies.
In dit gebied zijn de genen die coderen voor ribosomaal RNA gegroepeerd, getranscribeerd door RNA-polymerase I. In menselijk DNA worden deze genen aangetroffen in de satellieten van de volgende chromosomen: 13, 14, 15, 21 en 22. Dit zijn nucleolaire organisatoren.
De nucleolus is op zijn beurt gescheiden in drie afzonderlijke gebieden: fibrillaire centra, fibrillaire componenten en granulaire componenten..
Recente studies hebben steeds meer bewijs verzameld van mogelijke aanvullende functies van de nucleolus, niet alleen beperkt tot de synthese en assemblage van ribosomaal RNA.
Momenteel wordt aangenomen dat de nucleolus betrokken kan zijn bij de assemblage en synthese van verschillende eiwitten. Post-transcriptionele wijzigingen zijn ook aangetoond in deze nucleaire zone.
De nucleolus is ook betrokken bij regulerende functies. Een studie toonde aan hoe het verband hield met tumorsuppressoreiwitten.
De lichamen van Cajal (ook wel opgerolde lichamen) dragen deze naam ter ere van hun ontdekker, Santiago Ramón y Cajal. Deze onderzoeker observeerde deze bloedlichaampjes in neuronen in 1903.
Het zijn kleine structuren in de vorm van bolletjes en bestaan uit 1 tot 5 exemplaren per kern. Deze lichamen zijn erg complex met een vrij hoog aantal componenten, inclusief deze transcriptiefactoren en gerelateerde machinerie. splitsing.
Deze bolvormige structuren zijn gevonden in verschillende delen van de kern, aangezien het mobiele structuren zijn. Ze worden meestal in het nucleoplasma aangetroffen, hoewel ze in kankercellen in de nucleolus zijn aangetroffen.
Er zijn twee soorten Box-body's in de kern, ingedeeld naar hun grootte: groot en klein.
PML-lichamen (voor het acroniem in het Engels, promyelocytische leukemie) zijn kleine bolvormige subnucleaire gebieden met klinisch belang, aangezien ze in verband zijn gebracht met virale infecties en oncogenese.
Ze zijn bekend onder verschillende namen in de literatuur, zoals nucleair domein 10, Kremer-lichamen en oncogene PML-domeinen..
Een kern heeft 10 tot 30 van deze domeinen en ze hebben een diameter van 0,2 tot 1,0 µm. Onder zijn functies valt de regulatie van genen en RNA-synthese op..



Niemand heeft nog op dit artikel gereageerd.