
EEN operon bestaat uit een groep opeenvolgend geordende genen die elkaar reguleren, die coderen voor eiwitten die functioneel verwant zijn en die worden aangetroffen in het genoom van bacteriën en de 'voorouderlijke' genomen.
Dit reguleringsmechanisme werd in 1961 beschreven door F. Jacob en J. Monod, een feit dat hen in 1965 de Nobelprijs voor Fysiologie en Geneeskunde opleverde. Deze onderzoekers stelden de werking van operons voor en demonstreerden deze via de genen die coderen voor de enzymen die vereist zijn door Escherichia coli voor het gebruik van lactose.

Operons zijn verantwoordelijk voor het coördineren van de eiwitsynthese volgens de behoeften van elke cel, dat wil zeggen dat ze alleen tot expressie worden gebracht om eiwitten te genereren op het moment en op de exacte plaats waar ze nodig zijn..
De genen in operons zijn over het algemeen structurele genen, wat betekent dat ze coderen voor belangrijke enzymen die direct betrokken zijn bij metabole routes in de cel. Dit kan de synthese zijn van aminozuren, energie in de vorm van ATP, koolhydraten, etc..
Operons worden ook vaak aangetroffen in eukaryote organismen, maar in tegenstelling tot prokaryote organismen wordt in eukaryoten het gebied van het operon niet getranscribeerd als een enkel boodschapper-RNA-molecuul..
Artikel index
De eerste belangrijke stap vooruit met betrekking tot operons gemaakt door François Jacob en Jacques Monod was de focus op het probleem van "enzymatische aanpassing", dat erin bestond alleen een specifiek enzym te laten verschijnen als de cel in de aanwezigheid van een substraat was..
Zo'n reactie van cellen op substraten werd al jaren bij bacteriën waargenomen. De onderzoekers vroegen zich echter af hoe de cel precies bepaalde welk enzym het moest synthetiseren om dat substraat te metaboliseren..
Jacob en Monod merkten op dat bacteriële cellen, in aanwezigheid van galactose-achtige koolhydraten, 100 keer meer β-galactosidase produceerden dan onder normale omstandigheden. Dit enzym is verantwoordelijk voor het afbreken van β-galactosiden, zodat de cel ze metabolisch gebruikt.
Daarom noemden beide onderzoekers koolhydraten van het galactoside-type als "inductoren", aangezien ze verantwoordelijk waren voor het induceren van een toename van de synthese van β-galactosidase..
Evenzo vonden Jacob en Monod een genetisch gebied met drie genen die op een gecoördineerde manier werden gecontroleerd: het Z-gen, dat codeert voor het β-galactosidase-enzym; het Y-gen, dat codeert voor het enzym lactosepermease (galactosidetransport); en gen A, dat codeert voor het enzym transacetylase, dat ook essentieel is voor de assimilatie van galactosiden.
Door middel van daaropvolgende genetische analyses hebben Jacob en Monod alle aspecten van de genetische controle van het lactose-operon verduidelijkt en geconcludeerd dat het segment van de genen Z, Y en A een enkele genetische eenheid vormt met gecoördineerde expressie, wat ze definieerden als 'operon'..
Het operon-model werd voor het eerst nauwkeurig beschreven in 1965 door Jacob en Monod om de regulatie van genen te verklaren die worden getranscribeerd en vertaald voor de enzymen die nodig zijn in Escherichia coli om lactose als energiebron te metaboliseren.
Deze onderzoekers stelden voor dat de transcripten van het gen of van de reeks genen die opeenvolgend zijn gelokaliseerd, worden gereguleerd door twee elementen: 1) een regulerend gen of repressorgen 2) en een operatorgen of operatorsequentie.
Het operator-gen bevindt zich altijd naast het structurele gen (de structurele genen) waarvan de expressie verantwoordelijk is voor het reguleren, terwijl het repressorgen codeert voor een eiwit dat "repressor" wordt genoemd en dat bindt aan de operator en de transcriptie ervan verhindert..
Transcriptie wordt onderdrukt wanneer de repressor is gekoppeld aan het operator-gen. Op deze manier wordt de genetische expressie van de genen die coderen voor de enzymen die nodig zijn om lactose te assimileren, niet tot expressie gebracht en kan deze disaccharide daarom niet metaboliseren..
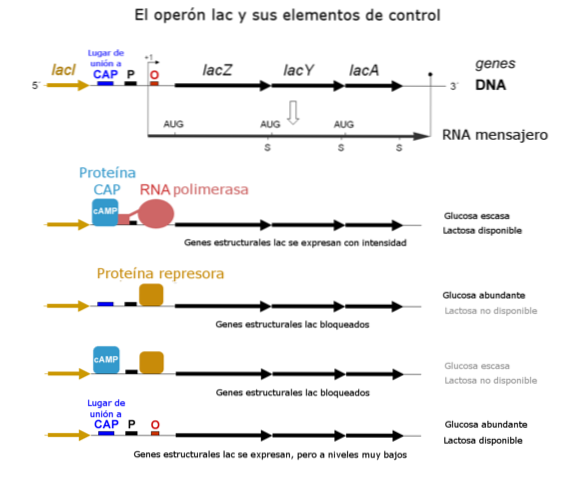
Momenteel is bekend dat de binding van de repressor aan de operator, met sterische mechanismen, verhindert dat het RNA-polymerase zich bindt aan de promotorplaats zodat het genen begint te transcriberen..
De promotorplaats is de "plaats" die RNA-polymerase herkent om genen te binden en te transcriberen. Omdat het niet kan binden, kan het geen van de genen in de sequentie transcriberen.
Het operator-gen ligt tussen een genetisch gebied van de sequentie dat bekend staat als de promotor en de structurele genen. Jacob en Monod hebben deze regio in hun tijd echter niet geïdentificeerd.
Het is momenteel bekend dat de volledige sequentie die het structurele gen of de structurele genen, de operator en de promotor omvat, in wezen een "operon" is..
Operons worden ingedeeld in slechts drie verschillende categorieën die afhankelijk zijn van de manier waarop ze worden gereguleerd, dat wil zeggen, sommige worden continu tot expressie gebracht (constitutief), andere hebben een specifiek molecuul of specifieke factor nodig om te activeren (induceerbaar) en andere worden continu tot expressie gebracht totdat de inductor is uitgedrukt (onderdrukbaar).
De drie soorten operons zijn:
Operons van dit type worden gereguleerd door moleculen in de omgeving, zoals aminozuren, suikers, metabolieten, enz. Deze moleculen staan bekend als inductoren. Als het molecuul dat als inductor fungeert, niet wordt gevonden, worden de operon-genen niet actief getranscribeerd.
In induceerbare operons bindt de vrije repressor zich aan de operator en voorkomt hij transcriptie van de genen die in het operon worden aangetroffen. Wanneer de inductor zich bindt aan de repressor, wordt een complex gevormd dat niet kan binden aan de repressor en daarom worden de genen van het operon vertaald.
Deze operons zijn afhankelijk van specifieke moleculen: onder andere aminozuren, suikers, cofactoren of transcriptiefactoren. Deze staan bekend als corepressoren en werken op een totaal tegenovergestelde manier als inductoren..
Alleen wanneer de corepressor zich bindt met de repressor, stopt de transcriptie en vindt dus de transcriptie van de genen in het operon niet plaats. Dan stopt de transcriptie van een onderdrukbaar operon alleen met de aanwezigheid van de corepressor.
Dit soort operons is niet gereguleerd. Ze worden constant actief getranscribeerd en in het geval van een mutatie die de sequentie van deze genen beïnvloedt, kan de levensduur van de cellen die ze bevatten worden aangetast en, in het algemeen, geprogrammeerde celdood teweegbrengen..
Het vroegste en meest bekende voorbeeld van de functie van een operon is de operon lac (lactose). Dit systeem is verantwoordelijk voor het omzetten van lactose, een disaccharide, in de monosacchariden glucose en galactose. Drie enzymen werken in dit proces:
- Β-galactosidase, verantwoordelijk voor het omzetten van lactose in glucose en galactose.
- Lactose permease, verantwoordelijk voor het transport van lactose van het extracellulaire medium naar het binnenste van de cel en
- Transketylase, dat tot het systeem behoort, maar een onbekende functie heeft
Het operon trp (tryptofaan) van Escherichia coli regelt de synthese van tryptofaan, met chorisminezuur als precursor. Binnen dit operon bevinden zich de genen voor vijf eiwitten die worden gebruikt voor de productie van drie enzymen:
- Het eerste enzym, gecodeerd door genen E en D, katalyseert de eerste twee reacties van de tryptofaanroute en staat bekend als anthranilaatsynthetase.
- Het tweede enzym is glycerolfosfaat en het katalyseert de stroomafwaartse stappen van anthranilaatsynthetase.
- Het derde en laatste enzym is tryptofaan synthetase, verantwoordelijk voor de productie van tryptofaan uit indool-glycerolfosfaat en serine (dit enzym is een product van genen B en A)



Niemand heeft nog op dit artikel gereageerd.