
De pachytene of paquinema is de derde fase van meiotische profase I; daarin wordt het recombinatieproces geverifieerd. In mitose is er één profase en in meiose twee: profase I en profase II.
Eerder, behalve voor profase II, werden de chromosomen gedupliceerd, wat elk aanleiding gaf tot een zusterchromatide. Maar alleen in de profase combineer ik homologen (duplicaten), waardoor ze bivalenten vormen.

De term paquiteno komt uit het Grieks en betekent "dikke draden". Deze "dikke draden" zijn de gepaarde homologe chromosomen die, na duplicatie, tetrads vormen. Dat zijn vier "draden", of snaren, die ervoor zorgen dat elk chromosoom dikker wordt.
Er zijn unieke aspecten van meiotische profase I die de inherente kenmerken van pachytene verklaren. Alleen in meiose profase I pachytene recombineren chromosomen.
Om dit te doen, wordt de herkenning en matching van de homologen geverifieerd. Net als bij mitose, moet er duplicatie van de chromatiden zijn. Maar pas in de meiose I pachytene worden banduitwisselingscomplexen gevormd, die we chiasmata noemen..
Daarin komt voor wat de recombinatiekracht van meiose definieert: de kruising tussen chromatiden van homologe chromosomen.
Het hele proces van DNA-uitwisseling is mogelijk dankzij het eerdere verschijnen van het synaptonemische complex. Dit multiproteïnecomplex maakt het mogelijk dat homologe chromosomen paren (synaps) en recombineren..
Artikel index
Het synaptonemische complex (CS) is het eiwitraamwerk dat een end-to-end-verbinding tussen homologe chromosomen mogelijk maakt. Het komt alleen voor tijdens de pachytene van meiose I, en is de fysieke basis van chromosomale paren. Met andere woorden, het is wat het mogelijk maakt dat chromosomen synaps en recombineren..
Het synaptonemische complex is sterk geconserveerd onder eukaryoten die meiose ondergaan. Daarom is het evolutionair erg oud en structureel en functioneel equivalent in alle levende wezens..
Het bestaat uit een centraal axiaal element en twee laterale elementen die worden herhaald als de tanden van een ritssluiting of sluiting..
Het synaptonemische complex wordt gevormd uit specifieke punten op de chromosomen tijdens zygoteen. Deze sites zijn collineair met die waar DNA-breuken optreden, waar synapsen en recombinatie in het pachytene zullen worden ervaren..
Tijdens de pachytene hebben we daarom een gesloten ritssluiting. In deze conformatie worden specifieke punten gedefinieerd waar DNA-banden aan het einde van de fase zullen worden uitgewisseld..
Het meiotische synaptonemische complex bevat veel structurele eiwitten die ook tijdens mitose worden aangetroffen. Deze omvatten topoisomerase II, condensines, cohesines, evenals met cohesine geassocieerde eiwitten..
Naast deze zijn er ook eiwitten die specifiek en uniek zijn voor meiose, samen met eiwitten van het recombinatiecomplex..
Deze eiwitten maken deel uit van het recombinosoom. Deze structuur groepeert alle eiwitten die nodig zijn voor recombinatie. Blijkbaar vormt het recombinosoom zich niet op de kruispunten, maar wordt het, al gevormd, naar hen toe gerekruteerd.
Chiasmen zijn de zichtbare morfologische structuren op chromosomen waar cross-overs voorkomen. Met andere woorden, de fysieke manifestatie van de uitwisseling van DNA-banden tussen twee homologe chromosomen. Chiasmen zijn de kenmerkende cytomorfologische kenmerken van pachytene.
Bij alle meiose moet er minstens één chiasma per chromosoom voorkomen. Dit betekent dat elke gameet recombinant is. Dankzij dit fenomeen konden de eerste genetische kaarten op basis van koppeling en recombinatie worden afgeleid en voorgesteld..
Aan de andere kant veroorzaakt het ontbreken van chiasmen, en dus van crossover, vervormingen op het niveau van chromosomale segregatie. Recombinatie tijdens pachytene fungeert dan als een kwaliteitscontrole van meiotische segregatie..
Evolutionair gezien ondergaan echter niet alle organismen recombinatie (bijvoorbeeld mannelijke fruitvliegen). In deze gevallen werken andere mechanismen van chromosomale segregatie die niet afhankelijk zijn van recombinatie..
Bij het verlaten van het zygoteen is het synaptonemische complex volledig gevormd. Dit wordt aangevuld door het genereren van dubbelbandige DNA-breuken waaruit cross-overs worden geverifieerd..
Dubbele DNA-breuken dwingen de cel om ze te repareren. Tijdens het DNA-herstel rekruteert de cel het recombinosoom. Banduitwisseling wordt gebruikt en als resultaat worden recombinante cellen verkregen.
Wanneer het synaptonemische complex volledig is gevormd, zou het pachytene beginnen.
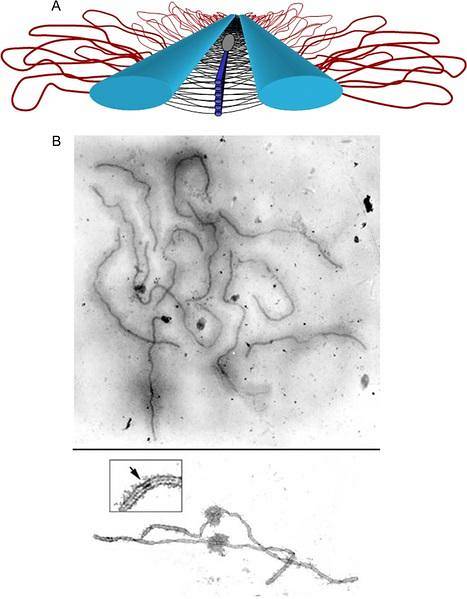
De bivalenten in synapsen in het pachytene werken in principe samen via het axiale element van het synaptonemische complex. Elke chromatide is georganiseerd in een lusorganisatie, waarvan de basis het centrale axiale element van het synaptonemische complex is..
Het axiale element van elke tegenhanger maakt contact met dat van de andere via de laterale elementen. De zusterchromatideassen zijn sterk verdicht en hun chromatinelussen komen naar buiten uit het centrale axiale element. Loopafstand (~ 20 per micrometer) is evolutionair geconserveerd voor alle soorten.
Richting het uiteinde van het pachytene zijn cross-overs duidelijk van enkele van de dubbelbandige DNA-breekplaatsen. Het verschijnen van de cross-overs markeert ook het begin van de ontrafeling van het synaptonemische complex..
Homologe chromosomen worden meer gecondenseerd (zien er meer individueel uit) en beginnen te scheiden, behalve in de chiasmata. Wanneer dit gebeurt, eindigt het pachytene en begint het diplotene..
De associatie tussen het recombinosoom en de assen van het synaptonemische complex blijft gedurende de synaps bestaan. Vooral bij recombinogene kruisingen naar het einde van het pachytene, of iets verder.



Niemand heeft nog op dit artikel gereageerd.