De RNA of RNA (ribonucleïnezuur) is een type nucleïnezuur dat aanwezig is in eukaryote en prokaryote organismen en in virussen. Het is een nucleotidepolymeer dat vier soorten stikstofhoudende basen in zijn structuur bevat: adenine, guanine, cytosine en uracil.
RNA wordt over het algemeen aangetroffen als een enkele band (behalve bij sommige virussen), op een lineaire manier of in een reeks complexe structuren. In feite heeft RNA een structurele dynamiek die niet wordt waargenomen in de dubbele DNA-helix. De verschillende soorten RNA hebben zeer uiteenlopende functies.
Ribosomale RNA's maken deel uit van ribosomen, de structuren die verantwoordelijk zijn voor de eiwitsynthese in cellen. Boodschapper-RNA's functioneren als tussenpersonen en transporteren genetische informatie naar het ribosoom, dat de boodschap vertaalt van een nucleotidesequentie naar een aminozuursequentie..
Transfer-RNA's zijn verantwoordelijk voor het activeren en overbrengen van de verschillende soorten aminozuren -20 in totaal naar de ribosomen. Er is een transfer-RNA-molecuul voor elk aminozuur dat de sequentie in boodschapper-RNA herkent.
Bovendien zijn er andere soorten RNA die niet direct betrokken zijn bij de eiwitsynthese en die deelnemen aan genregulatie..
Artikel index
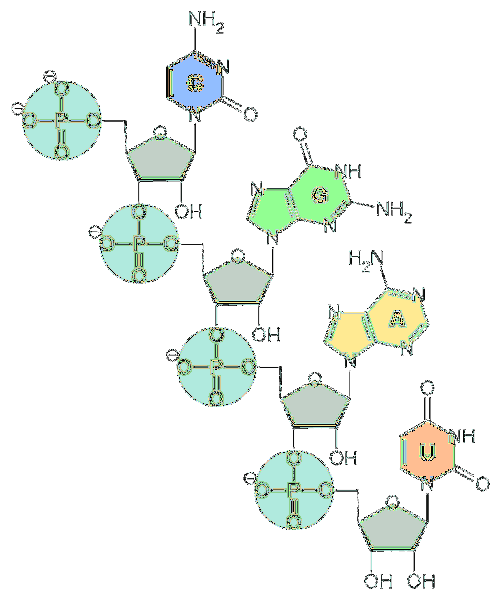
De fundamentele eenheden van RNA zijn nucleotiden. Elke nucleotide bestaat uit een stikstofbase (adenine, guanine, cytosine en uracil), een pentose en een fosfaatgroep..
Stikstofbasen zijn afgeleid van twee fundamentele verbindingen: pyrimidinen en purinen..
De basen afgeleid van purines zijn adenine en guanine en de basen afgeleid van pyrimidines zijn cytosine en uracil. Hoewel dit de meest voorkomende basen zijn, kunnen nucleïnezuren ook andere soorten basen hebben die minder vaak voorkomen..
Wat betreft pentose, het zijn eenheden van d-ribose. Om deze reden worden de nucleotiden waaruit RNA bestaat 'ribonucleotiden' genoemd..
Nucleotiden zijn met elkaar verbonden door chemische bindingen waarbij de fosfaatgroep betrokken is. Om ze te vormen, wordt de fosfaatgroep aan het 5'-uiteinde van een nucleotide gehecht aan de hydroxylgroep (-OH) aan het 3'-uiteinde van het volgende nucleotide, waardoor een fosfodiëster-achtige binding ontstaat..
Langs de nucleïnezuurketen hebben de fosfodiësterbindingen dezelfde oriëntatie. Daarom is er een polariteit van de streng, die onderscheid maakt tussen het 3'- en het 5'-uiteinde.
Volgens afspraak wordt de structuur van nucleïnezuren weergegeven door het 5'-uiteinde aan de linkerkant en het 3'-uiteinde aan de rechterkant.
Het RNA-product van DNA-transcriptie is een enkelstrengs band die naar rechts draait, in een spiraalvormige conformatie door het stapelen van basen. De interactie tussen purines is vanwege hun grootte veel groter dan de interactie tussen twee pyrimidines.
In RNA is het niet mogelijk om te spreken van een traditionele en referentie secundaire structuur, zoals de dubbele helix van DNA. De driedimensionale structuur van elk RNA-molecuul is uniek en complex, vergelijkbaar met die van eiwitten (logischerwijs kunnen we de structuur van eiwitten niet globaliseren).
Er zijn zwakke interacties die bijdragen aan de stabilisatie van RNA, met name base-stacking, waarbij de ringen zich op elkaar bevinden. Dit fenomeen draagt ook bij aan de stabiliteit van de DNA-helix..
Als het RNA-molecuul een complementaire sequentie vindt, kunnen ze koppelen en een dubbelstrengs structuur vormen die naar rechts draait. De overheersende vorm is type A; Wat betreft de Z-vormen, ze zijn alleen in het laboratorium aangetoond, terwijl de B-vorm niet is waargenomen.
Over het algemeen zijn er korte sequenties (zoals UUGG) die zich aan het einde van het RNA bevinden en de bijzonderheid hebben dat ze zich vormen lussen stal. Deze sequentie neemt deel aan het vouwen van de driedimensionale structuur van RNA.
Bovendien kunnen waterstofbruggen zich op andere plaatsen vormen dan de typische basenparen (AU en CG). Een van deze interacties vindt plaats tussen de 2'-OH van ribose met andere groepen.
Het ophelderen van de verschillende structuren die in RNA worden aangetroffen, heeft gediend om de vele functies van dit nucleïnezuur aan te tonen..
Er zijn twee klassen RNA: informatief en functioneel. De eerste groep omvat RNA's die deelnemen aan eiwitsynthese en functioneren als tussenpersonen in het proces; informatieve RNA's zijn boodschapper-RNA's.
Daarentegen geven RNA's die tot de tweede klasse behoren, de functionele, geen aanleiding tot een nieuw eiwitmolecuul en het RNA zelf is het eindproduct. Dit zijn transfer-RNA's en ribosomale RNA's..
In zoogdiercellen is 80% van het RNA ribosomaal RNA, 15% is transfer-RNA en slechts een klein deel komt overeen met boodschapper-RNA. Deze drie typen werken samen om de biosynthese van eiwitten te bereiken..
Er zijn onder andere ook kleine nucleaire RNA's, kleine cytoplasmatische RNA's en microRNA's. Elk van de belangrijkste typen wordt hieronder in detail beschreven:
In eukaryoten is DNA beperkt tot de kern, terwijl eiwitsynthese plaatsvindt in het cytoplasma van de cel, waar ribosomen worden gevonden. Vanwege deze ruimtelijke scheiding moet er een bemiddelaar zijn die de boodschap van de kern naar het cytoplasma overbrengt en dat molecuul is het boodschapper-RNA..
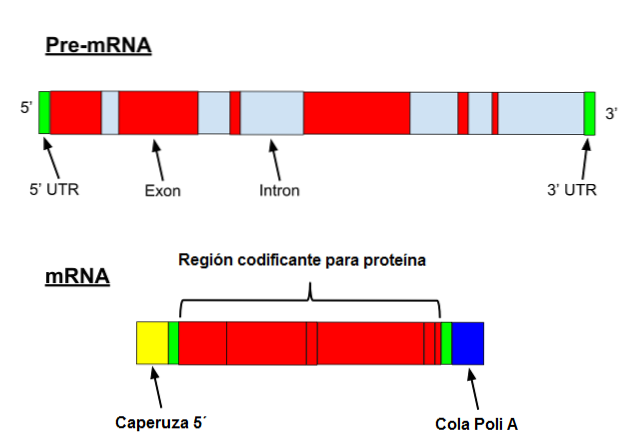
Messenger RNA, afgekort mRNA, is een intermediair molecuul dat de informatie bevat die in DNA wordt gecodeerd en dat een aminozuursequentie specificeert die aanleiding geeft tot een functioneel eiwit..
De term boodschapper-RNA werd in 1961 voorgesteld door François Jacob en Jacques Monod om het deel van het RNA te beschrijven dat de boodschap van DNA naar ribosomen overbracht..
Het proces van synthese van een mRNA uit de DNA-streng staat bekend als transcriptie en vindt differentieel plaats tussen prokaryoten en eukaryoten..
Genexpressie wordt bepaald door verschillende factoren en hangt af van de behoeften van elke cel. Transcriptie is onderverdeeld in drie fasen: initiatie, verlenging en beëindiging..
Het DNA-replicatieproces, dat plaatsvindt in elke celdeling, kopieert het hele chromosoom. Het transcriptieproces is echter veel selectiever, het behandelt alleen de verwerking van specifieke segmenten van de DNA-streng en vereist geen primer..
Aan Escherichia coli -de best bestudeerde bacterie in de levenswetenschappen - transcriptie begint met het afwikkelen van de dubbele DNA-helix en de transcriptielus wordt gevormd. Het enzym RNA-polymerase is verantwoordelijk voor het synthetiseren van RNA en naarmate de transcriptie vordert, keert de DNA-streng terug naar zijn oorspronkelijke vorm.
Transcriptie wordt niet gestart op willekeurige plaatsen op het DNA-molecuul; er zijn gespecialiseerde sites voor dit fenomeen, promotors genaamd. Aan E coli RNA-polymerase is een paar basenparen boven het doelgebied gekoppeld.
De sequenties waaraan transcriptiefactoren zijn gekoppeld, zijn behoorlijk geconserveerd tussen verschillende soorten. Een van de bekendste promotersequenties is de TATA-box.
Bij verlenging voegt het RNA-polymerase-enzym nieuwe nucleotiden toe aan het 3'-OH-uiteinde, in de richting van 5 'naar 3'. De hydroxylgroep werkt als een nucleofiel en valt het alfafosfaat van het toe te voegen nucleotide aan. Bij deze reactie komt een pyrofosfaat vrij.
Slechts één van de DNA-strengen wordt gebruikt om het boodschapper-RNA te synthetiseren, dat wordt gekopieerd in de 3 'naar 5' richting (de antiparallelle vorm van de nieuwe RNA-streng). Het toe te voegen nucleotide moet voldoen aan basenparing: U-paren met A en G met C.
RNA-polymerase stopt het proces wanneer het regio's vindt die rijk zijn aan cytosine en guanine. Uiteindelijk wordt het nieuwe boodschapper-RNA-molecuul gescheiden van het complex.
In prokaryoten kan een boodschapper-RNA-molecuul coderen voor meer dan één eiwit.
Wanneer een mRNA exclusief codeert voor een eiwit of polypeptide, wordt het monocistronisch mRNA genoemd, maar als het codeert voor meer dan één eiwitproduct, is het mRNA polycistronisch (merk op dat in deze context de term cistron verwijst naar het gen).
In eukaryote organismen is de overgrote meerderheid van mRNA's monocistronisch en de transcriptionele machinerie is veel complexer in deze lijn van organismen. Ze worden gekenmerkt door het feit dat ze drie RNA-polymerasen hebben, aangeduid met I, II en III, elk met specifieke functies.
De I is verantwoordelijk voor het synthetiseren van de pre-rRNA's, de II synthetiseert de boodschapper-RNA's en enkele speciale RNA's. Ten slotte is III verantwoordelijk voor transfer-RNA's, ribosomale 5S en andere kleine RNA's..
Messenger RNA ondergaat een reeks specifieke wijzigingen in eukaryoten. De eerste betreft het toevoegen van een "dop" aan het 5'-uiteinde. Chemisch gezien is de dop een 7-methylguanosineresidu dat aan het uiteinde is bevestigd door een 5 ', 5'-trifosfaatbinding..
De functie van deze zone is om RNA te beschermen tegen mogelijke afbraak door ribonucleasen (enzymen die RNA afbreken in kleinere componenten).
Bovendien vindt verwijdering van het 3'-uiteinde plaats en worden 80 tot 250 adenineresiduen toegevoegd. Deze structuur staat bekend als de polyA "staart" en dient als bindingsplaats voor verschillende eiwitten. Wanneer een prokaryoot een polyA-staart verwerft, heeft het de neiging de afbraak ervan te stimuleren..
Aan de andere kant wordt deze boodschapper getranscribeerd met introns. Introns zijn DNA-sequenties die geen deel uitmaken van het gen, maar die sequentie "onderbreken". Introns worden niet vertaald en moeten daarom uit de messenger worden verwijderd.
De meeste genen van gewervelde dieren hebben introns, met uitzondering van genen die coderen voor histonen. Evenzo kan het aantal introns in een gen variëren van enkele tot tientallen..
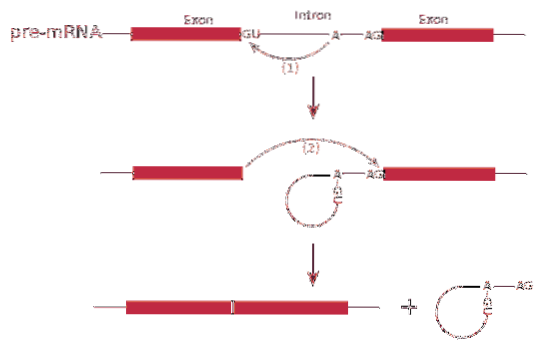
De splicing RNA of splitsingsproces omvat de verwijdering van introns in boodschapper-RNA.
Sommige introns die in nucleaire of mitochondriale genen worden aangetroffen, kunnen het proces van splitsing zonder de hulp van enzymen of ATP. In plaats daarvan wordt het proces uitgevoerd door omesteringsreacties. Dit mechanisme werd ontdekt in de trilharenprotozoa Tetrahymena thermophila.
Daarentegen is er een andere groep boodschappers die niet in staat zijn hun eigen te bemiddelen splitsing, dus hebben ze extra machines nodig. Tot deze groep behoort een vrij groot aantal nucleaire genen..
Het proces van splitsing het wordt gemedieerd door een eiwitcomplex dat een spliceosoom of splitsingscomplex wordt genoemd. Het systeem bestaat uit gespecialiseerde RNA-complexen die kleine nucleaire ribonucleoproteïnen (RNP) worden genoemd..
Er zijn vijf soorten RNP: U1, U2, U4, U5 en U6, die in de kern worden aangetroffen en het proces van splitsing.
De splitsing kan meer dan één type eiwit produceren - dit staat bekend als splitsing alternatief, aangezien de exons differentieel zijn gerangschikt, waardoor variëteiten van boodschapper-RNA's ontstaan.
Ribosomaal RNA, afgekort rRNA, wordt aangetroffen in ribosomen en neemt deel aan de biosynthese van eiwitten. Daarom is het een essentieel onderdeel van alle cellen.
Ribosomaal RNA associeert zich met eiwitmoleculen (ongeveer 100) waardoor ribosomale presubeenheden ontstaan. Ze worden geclassificeerd op basis van hun sedimentatiecoëfficiënt, aangeduid met de letter S voor Svedberg-eenheden..
Een ribosoom bestaat uit twee delen: de hoofdsubeenheid en de kleinere subeenheid. Beide subeenheden verschillen tussen prokaryoten en eukaryoten in termen van de sedimentatiecoëfficiënt..
Prokaryoten hebben een grote 50S-subeenheid en een kleine 30S-subeenheid, terwijl in eukaryoten de grote subeenheid 60S is en de kleine 40S.
De genen die coderen voor ribosomale RNA's bevinden zich in de nucleolus, een bepaald gebied van de kern dat niet wordt begrensd door een membraan. Ribosomale RNA's worden in dit gebied getranscribeerd door RNA-polymerase I.
In cellen die grote hoeveelheden eiwitten aanmaken; de nucleolus is een prominente structuur. Wanneer de cel in kwestie echter niet een groot aantal eiwitproducten nodig heeft, is de nucleolus een bijna onmerkbare structuur.
De 60S grote ribosomale subeenheid is geassocieerd met de 28S- en 5.8S-fragmenten. Wat betreft de kleine subeenheid (40S), deze wordt geassocieerd met 18S.
In hogere eukaryoten wordt pre-rRNA gecodeerd in een 45S-transcriptie-eenheid, die RNA-polymerase I omvat. Dit transcript wordt verwerkt tot rijpe 28S-, 18S- en 5.8S-ribosomale RNA's..
Naarmate de synthese vordert, associeert het pre-rRNA zich met verschillende eiwitten en vormt het ribonucleoproteïnedeeltjes. Het ondergaat een reeks opeenvolgende modificaties, waaronder methylering van de 2'-OH-groep van ribose en de omzetting van uridineresiduen in pseudouridine..
Het gebied waar deze veranderingen plaatsvinden, wordt gecontroleerd door meer dan 150 kleine nucleolaire RNA-moleculen, die het vermogen hebben om te koppelen aan pre-rRNA..
In tegenstelling tot de rest van de pre-rRNA's, wordt 5S getranscribeerd door RNA-polymerase III in het nucleoplasma en niet in de nucleolus. Na te zijn gesynthetiseerd, wordt het naar de nucleolus gebracht om te assembleren met de 28S en 5.8S, waardoor de ribosomale eenheden worden gevormd.
Aan het einde van het assemblageproces worden de subeenheden via de nucleaire poriën overgebracht naar het cytoplasma..
Het kan gebeuren dat een molecuul boodschapper-RNA tegelijkertijd meerdere eiwitten aanmaakt, die aan meer dan één ribosoom binden. Naarmate het vertaalproces vordert, komt het einde van de boodschapper vrij en kan het worden opgenomen door een ander ribosoom, waarmee een nieuwe synthese wordt gestart..
Om deze reden is het gebruikelijk om ribosomen gegroepeerd (tussen 3 en 10) te vinden in een enkel molecuul boodschapper-RNA, en deze groep wordt polyribosoom genoemd..
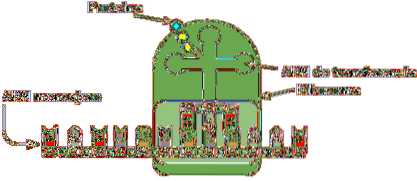
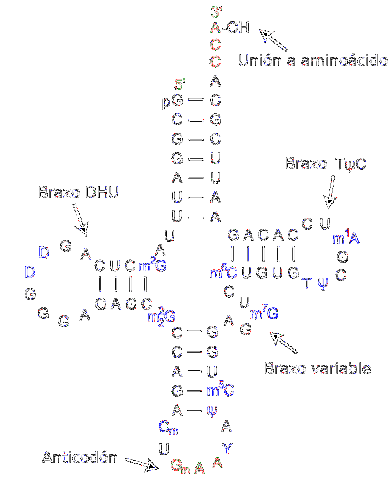
Transfer RNA is verantwoordelijk voor de overdracht van aminozuren naarmate het eiwitsyntheseproces vordert. Ze bestaan uit ongeveer 80 nucleotiden (vergeleken met boodschapper-RNA is het een "klein" molecuul).
De structuur heeft plooien en kruisen die doen denken aan een driearmige klaver. Een adenylring bevindt zich aan een van de uiteinden, waar de hydroxylgroep van ribose de vereniging met het te transporteren aminozuur bemiddelt.
De verschillende transfer-RNA's worden exclusief gecombineerd met een van de twintig aminozuren waaruit eiwitten bestaan; met andere woorden, het is het voertuig dat de bouwstenen van eiwitten draagt. Het transfer-RNA-complex samen met het aminozuur wordt aminoacyl-tRNA genoemd.
Bovendien herkent elk transfer-RNA in het translatieproces - dat plaatsvindt dankzij ribosomen - een specifiek codon in het boodschapper-RNA. Wanneer het wordt herkend, wordt het overeenkomstige aminozuur vrijgegeven en wordt het onderdeel van het gesynthetiseerde peptide.
Om het type aminozuur dat moet worden afgegeven te herkennen, heeft het RNA een "anticodon" in het middelste gebied van het molecuul. Dit anticodon is in staat om waterstofbruggen te vormen met de complementaire basen die aanwezig zijn in boodschapper-DNA..
MicroRNA's of miRNA's zijn een soort kort, enkelstrengs RNA, tussen 21 en 23 nucleotiden, waarvan de functie is om de expressie van genen te reguleren. Omdat het niet in proteïne wordt vertaald, wordt het vaak niet-coderend RNA genoemd..
Net als andere soorten RNA is microRNA-verwerking complex en omvat een aantal eiwitten.
MicroRNA's ontstaan uit langere voorlopers, mi-pri-RNA's genaamd, afgeleid van het eerste transcript van het gen. In de kern van de cel worden deze voorlopers gemodificeerd in het microprocessorcomplex en het resultaat is een pre-miRNA..
Pre-miRNA's zijn haarspelden met 70 nucleotiden die hun verwerking in het cytoplasma voortzetten door een enzym genaamd Dicer, dat het RNA-geïnduceerde silencing-complex (RISC) assembleert en tenslotte wordt het miRNA gesynthetiseerd..
Deze RNA's zijn in staat de expressie van genen te reguleren, aangezien ze complementair zijn aan specifieke boodschapper-RNA's. Door te paren met hun doelwit, kunnen de miRNA's de boodschapper onderdrukken of zelfs degraderen. Bijgevolg kan het ribosoom genoemd transcript niet vertalen..
Een bepaald type microRNA zijn kleine interfererende RNA's (siRNA's), ook wel silencing-RNA's genoemd. Het zijn korte RNA's, tussen de 20 en 25 nucleotiden, die de expressie van bepaalde genen belemmeren.
Het zijn veelbelovende instrumenten voor onderzoek, omdat ze het mogelijk maken om een gen van interesse het zwijgen op te leggen en zo de mogelijke functie ervan te bestuderen..
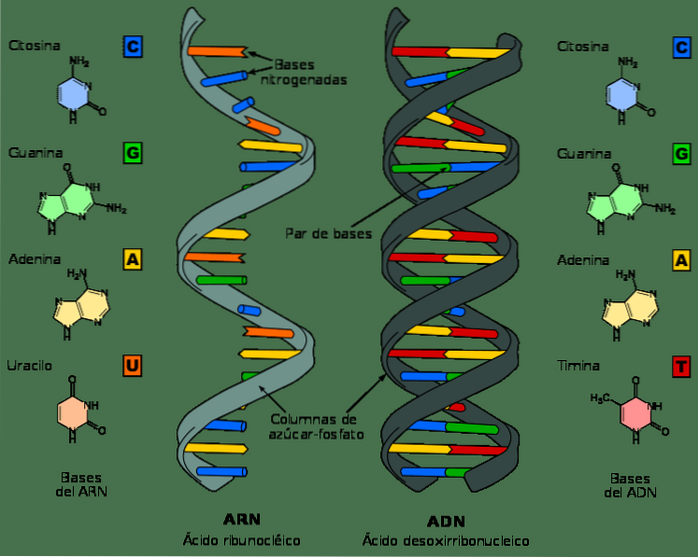
Hoewel DNA en RNA nucleïnezuren zijn en op het eerste gezicht erg op elkaar kunnen lijken, verschillen ze in verschillende van hun chemische en structurele eigenschappen. DNA is een molecuul met een dubbele band, terwijl RNA een enkele band is.
Als gevolg hiervan is RNA een veelzijdiger molecuul en kan het een grote verscheidenheid aan driedimensionale vormen aannemen. Bepaalde virussen hebben echter dubbelband-RNA in hun genetisch materiaal..
In RNA-nucleotiden is het suikermolecuul een ribose, terwijl het in DNA een deoxyribose is, die alleen verschilt in de aanwezigheid van een zuurstofatoom.
De fosfodiësterbinding in de ruggengraat van DNA en RNA ondergaat een langzaam hydrolyseproces zonder de aanwezigheid van enzymen. Onder alkalische omstandigheden hydrolyseert RNA snel - dankzij de extra hydroxylgroep - terwijl DNA dat niet doet.
Evenzo zijn de stikstofbasen waaruit nucleotiden in DNA bestaan guanine, adenine, thymine en cytosine; in tegenstelling daarmee wordt in RNA thymine vervangen door uracil. Uracil kan paren met adenine, op dezelfde manier als thymine in DNA.
RNA is het enige bekende molecuul dat informatie kan opslaan en tegelijkertijd chemische reacties kan katalyseren; Om deze reden stellen verschillende auteurs dat het RNA-molecuul cruciaal was bij het ontstaan van leven. Verrassend genoeg zijn ribosoomsubstraten andere RNA-moleculen.
De ontdekking van ribozymen leidde tot de biochemische herdefinitie van "enzym" - aangezien de term voorheen uitsluitend werd gebruikt voor eiwitten met katalytische activiteit - en hielp een scenario te ondersteunen waarin de eerste levensvormen alleen RNA als genetisch materiaal gebruikten..



Niemand heeft nog op dit artikel gereageerd.