De manier van pentosen Fosfaat, ook bekend als de omleiding van hexosemonofosfaat, is een fundamentele metabolische route waarvan het eindproduct ribosen is, noodzakelijk voor nucleotide- en nucleïnezuursyntheseroutes, zoals DNA, RNA, ATP, NADH, FAD en co-enzym A.
Het produceert ook NADPH (nicotinamide adenine dinucleotide fosfaat), gebruikt bij verschillende enzymatische reacties. Dit pad is zeer dynamisch en kan zijn producten aanpassen aan de momentane behoeften van de cellen..

ATP (adenosinetrifosfaat) wordt beschouwd als de "energiemunt" van de cel, omdat de hydrolyse ervan kan worden gekoppeld aan een breed scala aan biochemische reacties..
Op dezelfde manier is NADPH een essentiële tweede energiemunt voor onder andere de reductieve synthese van vetzuren, cholesterolsynthese, neurotransmittersynthese, fotosynthese en ontgiftingsreacties..
Hoewel NADPH en NADH qua structuur vergelijkbaar zijn, kunnen ze niet door elkaar worden gebruikt in biochemische reacties. NADPH neemt deel aan het gebruik van vrije energie bij de oxidatie van bepaalde metabolieten voor reductieve biosynthese.
NADH daarentegen is betrokken bij het gebruik van vrije energie uit de oxidatie van metabolieten om ATP te synthetiseren..
Artikel index
De aanwijzingen voor het bestaan van dit pad begonnen in 1930 dankzij de onderzoeker Otto Warburg, aan wie de ontdekking van NADP wordt toegeschreven+.
Bepaalde waarnemingen maakten de ontdekking van de route mogelijk, met name de voortzetting van de ademhaling in aanwezigheid van glycolyseremmers, zoals het fluoride-ion..
Vervolgens beschreven de wetenschappers Frank Dickens, Bernard Horecker, Fritz Lipmann en Efraim Racker in 1950 de pentosefosfaatroute.
Weefsels die betrokken zijn bij de synthese van cholesterol en vetzuren, zoals de borstklieren, vetweefsel en nieren, hebben hoge concentraties pentosefosfaat-enzymen..
De lever is ook een belangrijk weefsel voor deze route: ongeveer 30% van de glucose-oxidatie in dit weefsel vindt plaats dankzij de enzymen van de pentosefosfaatroute..
De pentosefosfaatroute is verantwoordelijk voor het in stand houden van koolstofhomeostase in de cel. Evenzo synthetiseert de route de voorlopers van nucleotiden en moleculen die betrokken zijn bij de synthese van aminozuren (de bouwstenen van peptiden en eiwitten)..
Het is de belangrijkste bron van reducerend vermogen voor enzymatische reacties. Bovendien levert het de moleculen die nodig zijn voor anabole reacties en voor afweerprocessen tegen oxidatieve stress. De laatste fase van het pad is cruciaal in redoxprocessen onder stresssituaties.
De pentosefosfaatroute bestaat uit twee fasen in het celcytosol: een oxidatieve, die NADPH genereert met de oxidatie van glucose-6-fosfaat tot ribose-5-fosfaat; en een niet-oxidatieve, die de onderlinge omzetting van suikers van drie, vier, vijf, zes en zeven koolstofatomen inhoudt.
Deze route presenteert reacties die worden gedeeld met de Calvin-cyclus en met de Entner-Doudoroff-route, die een alternatief is voor glycolyse..
De oxidatieve fase begint met de dehydrogenering van het glucose-6-fosfaatmolecuul op koolstof 1. Deze reactie wordt gekatalyseerd door het enzym glucose-6-fosfaatdehydrogenase, dat een hoge specificiteit heeft voor NADP.+.
Het product van deze reactie is 6-fosfonoglucono-δ-lacton. Dit product wordt vervolgens gehydrolyseerd door het enzym lactonase om 6-fosfogluconaat te geven. Deze laatste verbinding wordt opgenomen door het enzym 6-fosfogluconaat dehydrogenase en wordt ribulose 5-fosfaat.
Het enzym fosfopentose-isomerase katalyseert de laatste stap van de oxidatieve fase, die de synthese van ribose 5-fosfaat omvat door de isomerisatie van ribulose 5-fosfaat.
Deze reeks reacties produceert twee moleculen NADPH en één molecuul ribose 5-fosfaat voor elk molecuul glucose 6-fosfaat dat deze enzymatische route binnengaat..
In sommige cellen zijn de vereisten voor NADPH groter dan die voor ribose 5-fosfaat. Daarom nemen de enzymen transketolase en transaldolase ribose 5-fosfaat en zetten dit om in glyceraldehyde 3-fosfaat en fructose 6-fosfaat, waardoor plaats wordt gemaakt voor de niet-oxidatieve fase. Deze laatste twee verbindingen kunnen de glycolytische route binnendringen.
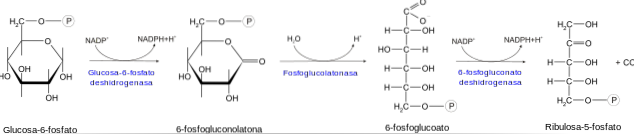
De fase begint met een epimerisatiereactie die wordt gekatalyseerd door het enzym pentose-5-fosfaatepimerase. Ribulose-5-fosfaat wordt door dit enzym opgenomen en omgezet in xylulose-5-fosfaat.
Het product wordt opgenomen door het enzym transketolase dat samenwerkt met het co-enzym thiamine pyrofosfaat (TTP), dat de doorgang van xylulose-5-fosfaat naar ribose-5-fosfaat katalyseert. Met de overdracht van ketose naar aldose worden glyceraldehyde-3-fosfaat en sedoheptulose-7-fosfaat geproduceerd.
Het enzym transaldolase draagt vervolgens het C3 over van het sedoheptulose-7-fosfaatmolecuul naar glyceraldehyde-3-fosfaat, waarbij een suiker met vier koolstofatomen (erythrose-4-fosfaat) en een suiker met zes koolstofatomen (fructose-6-fosfaat) wordt geproduceerd. Deze producten kunnen de glycolytische route voeden.
Het enzym transketosala zorgt weer voor de overdracht van een C2 van xylulose-5-fosfaat naar erythrose-4-fosfaat, wat resulteert in fructose-6-fosfaat en glyceraldehyde-3-fosfaat. Net als in de vorige stap kunnen deze producten glycolyse binnendringen.
Deze tweede fase verbindt de paden die NADPH genereren met degenen die verantwoordelijk zijn voor het synthetiseren van ATP en NADH. Bovendien kunnen de producten fructose-6-fosfaat en glyceraldehyde-3-fosfaat gluconeogenese ingaan..
Er zijn verschillende pathologieën gerelateerd aan de pentosefosfaatroute, tussen deze neuromusculaire ziekten en verschillende soorten kanker.
De meeste klinische onderzoeken zijn gericht op het kwantificeren van de activiteit van glucose-6-fosfaatdehydrogenase, omdat dit het belangrijkste enzym is dat verantwoordelijk is voor het reguleren van de route..
In bloedcellen die behoren tot personen die vatbaar zijn voor anemie, vertonen ze een lage enzymatische activiteit van glucose-6-fosfaatdehydrogenase. Daarentegen vertonen cellijnen die verband houden met carcinomen in het strottenhoofd een hoge enzymactiviteit..
NADPH is betrokken bij de productie van glutathion, een belangrijk peptidemolecuul ter bescherming tegen reactieve zuurstofsoorten, betrokken bij oxidatieve stress.
Verschillende soorten kanker leiden tot de activering van de pentose-route en het wordt geassocieerd met processen van metastase, angiogenese en reacties op chemotherapie en radiotherapiebehandelingen..
Aan de andere kant ontstaat chronische granulomateuze ziekte wanneer er een tekort is in de productie van NADPH.


Niemand heeft nog op dit artikel gereageerd.